In this tutorial, we have focused on ‘Transportation in Plants and Means of Transport’ based on three topics: ‘Water absorption and transport’, ‘Mineral transport’, and ‘Phloem transport’.
— Plants like other living organisms, i.e., humans and animals, also need to transport various types of substances such as gases, minerals, water, hormones, food, and organic solutes to short as well as long distances. All this transport takes place in absence of interstitial fluid as well as a regular circulation system.
{kind=link}
— The long distance transport that occurs through vascular tissues, i.e., xylem and phloem, is called translocation. It occurs through mass flow. The direction of translocation is essentially unidirectional in case of water (from root to stem, leaves, flowers, and fruits). It is multidirectional in case of minerals and organic solutes. Organic compounds synthesized in the photosynthetic leaves are exported to all other parts of the plant including storage organs. From the storage organs, they are later re-exported.
Hormones or plant growth regulators and other chemical stimuli are also transported, though in very small amounts, sometimes in a strictly polarized or unidirectional manner, from where they are synthesized, to other parts.
► Read More: Aerobic and Anaerobic Respiration in Plants ► Read More: What is ATP in Biology: Structure, Phosphorylation, Function ► Read More: Example of Biofertilizers, Organic & Chemical Fertilizers ► Read More: What is Transpiration in Plants?
TABLE OF CONTENTS
MEANS OF TRANSPORT
Passage of materials into and out of the cells is carried out by the following methods – diffusion, facilitated diffusion, active transport.
{kind=link}
{kind=link}
Diffusion is the movement of materials along the concentration gradient, i.e., from the region of higher concentration to the region of lower concentration. It is slow and passive, i.e., no energy expenditure takes place.
Diffusion is very important to plants, as it is the only means for gaseous movement within the plant body. The diffusing molecules or ions exert a pressure called diffusion pressure (DP). This can be defined as the potential ability of a molecule or ion (be it a solid, liquid, or gas) to diffuse from a region of its greatest concentration to the region of its lower concentration. The diffusion pressure is directly proportional to the concentration or the number of diffusing particles, i.e., the greater the concentration of diffusing particles in the system, their diffusion pressure will also be greater and vice versa.
Pure water has the maximum diffusion pressure. When solute particles are added to it, diffusion pressure gets lowered. The fall in the diffusion pressure of a solution in relation to pure water is called diffusion pressure deficit (DPD). Meyer, in 1938, coined the term diffusion pressure deficit (DPD). Due to the presence of diffusion pressure deficit, the solution tends to make up the deficit by absorbing water. Hence, it is also called suction pressure (SP).
{kind=link}
In case, two solutions OP1 and OP2 are separated by a semipermeable membrane, the DPD will be equal to the difference in the osmotic pressure of two solutions (OP1 and OP2) minus wall pressure or turgor pressure, i.e.,
DPD = (OP1 – OP2) – WP or TP
Similarly, if a cell is placed in a solution, its DPD will be the difference in the osmotic pressure of cell sap and external solution minus wall pressure.
DPD determines the speed of absorption or loss of water by the cell or system. The unit of DPD is atm or bar.
{kind=link}
► Factors influencing the Rate of Diffusion
Diffusion rates are affected by the gradient of concentration, the permeability of the membrane separating them, temperature, and pressure.
01. Temperature: Increase in temperature leads to an increase in the rate of diffusion because it increases the kinetic energy of diffusing particles.
02. Density of Diffusing Substance: The rate of diffusion is inversely proportional to the square root of the density of the diffusing substance. Larger the molecules, slower will be the rate of diffusion. This is also called Graham’s Law of Diffusion.
{kind=link}
03. Density of Diffusing Medium: Higher density of diffusing medium lowers the rate of diffusion. Gas molecules diffuse more rapidly in a vacuum than in air. Thus, the rate of diffusion is inversely proportional to the density of diffusing medium.
04. Diffusion Pressure Gradient: Diffusion pressure gradient develops by the concentration of diffusing substance. The higher is the difference in the concentration of diffusing substances in two solutions, the higher will be diffusion pressure gradient and rapid will be the rate of diffusion. The rate of diffusion is directly proportional to the diffusion pressure gradient at the two ends of a system and inversely proportional to the distance between the two.
{kind=link}
01. Exchange of Gases: Exchange of gases (CO2 and O2), between the plant interior and outside air, occurs through diffusion via stomata.
02 Transpiration: Loss of water vapours from the aerial parts of plants, i.e., transpiration, involves the process of diffusion.
03. Absorption: During passive intake of salts, the ions are absorbed by diffusion.
04. Translocation: It also helps in the translocation of food material.
05: Pollination: Aroma of flowers is due to the diffusion of volatile aromatic compounds and helps in attracting the pollinating organisms to carry out pollination.
06. Intracellular Transport: Various ions and substances rapidly spread through the cytoplasm due to diffusion.
07. Distribution of Water and Solutes: Diffusion brings about the distribution of water and solutes inside the plants. Additionally, diffusion keeps the cell walls of the internal plant tissues moist.
Facilitated Diffusion
The passive absorption of hydrophilic substances, mediated by a carrier, is called facilitated diffusion. It takes along a concentration gradient and no energy is utilized. The movement is facilitated through fixed membrane transport proteins.
Usually, substances that are lipid soluble, can easily pass directly through the cell membrane, as it is mainly made of lipids. The hydrophilic solutes and large charged polar molecules find it difficult to pass through the membrane, so their movement has to be facilitated. Their movement through the membrane is assisted by transport proteins.
The transport proteins allow the passage of selected ions and other polar molecules. They may be of two types – carrier proteins and channel proteins.
– Carrier proteins (also called carriers, transporters) bind the particular solute and transport it to the other side of the membrane. Carriers are highly selective for a particular substrate to be transported. A conformational change in the protein is required to transport individual molecules or ions hence the rate of transport is slow.
{kind=link}
● A molecule, when moves across the membrane through carrier protein independently of other molecules, the process is
called uniport.
● Some carrier proteins allow transport only if two types of molecules move together, this process is called cotransport. In cotransport, when both the molecules cross the membrane in the same direction at the same time, it is called symport and when both the molecules move in the opposite directions, the process is called antiport.
– Channel proteins are transmembrane proteins that function as selective pores through which molecules or ions can diffuse across the membrane. The size of the pore and density of surface charges on its interior lining determine its transport specificity. They allow the diffusion of water-soluble solutes of an appropriate size. Channels are not open all the time. They have structures called gates that open and close the pore in response to external signals (voltage changes, hormone binding, or light).
They are of two types – ion channels and porins.
● Ion channels allow passage of specific ions, e.g., K+ channels, during nerve conduction. Ion channels may open in different voltage ranges or in response to different signals which may include K+ or Ca2+ concentrations, pH, kinases, etc. This specificity enables the transport of each ion to be fine-tuned to the prevailing conditions.
● Porins are large protein pores in the outer membranes of the plastids, mitochondria, and some bacteria that allow even small sized proteins to pass through. For example, aquaporins, are water channels through which a massive amount of water diffuses into the cell.
{kind=link}
Active Transport
Active transport involves movable carrier proteins (not by channel proteins) called pumps which employ ATP energy for transport across the membrane. It is an uphill transport, i.e., pumps the solutes against the concentration gradient and is faster than passive transport. The rate of active transport reaches the maximum when all the protein pumps are used in transport, which is called the saturation effect.
Like enzymes, carrier proteins are very specific for transporting molecules and are also sensitive to inhibitors.
■ Comparison of Different Transport Processes
Proteins in the membrane are responsible for facilitated diffusion and active transport. Thus, they show common characteristics of being highly selective. They are liable to saturate, respond to inhibitors and are under hormonal control. But, diffusion, whether simple or facilitated takes part only along a gradient and does not use energy.
TABLE: Comparison between Different Transport Processes
Property Simple Diffusion Facilitated Diffusion Active Transport 1. Type of molecule required Lipids Proteins Proteins 2. Requires special membrane proteins No Carrier proteins and channel proteins Carrier proteins 3. Specificity Nonspecific Specific Specific 4. Transport saturates No Yes Yes 5. Uphill transport No No Yes 6. Requires ATP energy No No Yes 7. Direction of movement Along concentration gradient Along concentration gradient Against concentration gradient 8. Response to protein inhibitions No Yes Yes
[/su_note]
PLANT- WATER RELATIONSHIP
Water is essential for all physiological activities of the plant. It provides a medium in which substances get dissolved and undergo various types of reactions in all living organisms. To comprehend plant-water relation, an understanding of the following standard terms is necessary.
Membrane Permeability
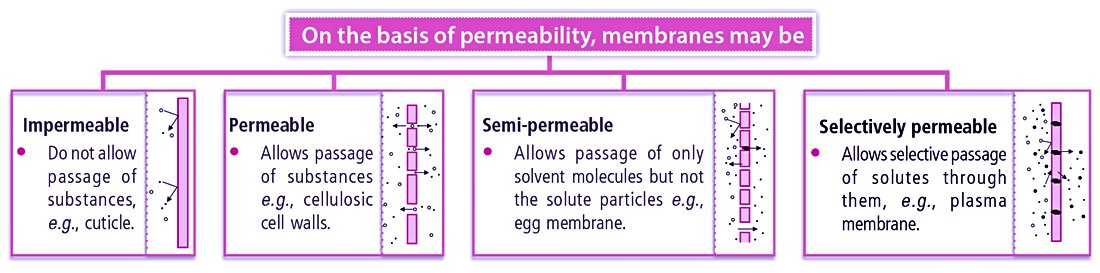
When a membrane allows the passage of a substance through it, it is said to be permeable and this property of the membrane is termed permeability. Membrane permeability is the ability of a membrane to allow the passage of gases, liquids, solutes, or dissolved substances through it. Depending upon the membrane composition and the chemical nature of the solute, a membrane can be impermeable, permeable, semipermeable, or selectively or differentially permeable.
01. Impermeable: Such membranes do not permit diffusion of water and solutes into and out of the cell, e.g., cuticle of plant cell walls, suberised wall of cork cells.
02. Permeable: Permeable membranes allow free movement of water and solutes into and out of the cell, e.g., cell wall in plant cells.
03. Semipermeable: Such membranes allow passage of only solvent molecules, but not the solute particles, e.g., egg membrane, parchment membrane.
04. Selectively or Differentially Permeable: Such membranes have protein-lined channels called aquaporins for the passage of water and ion channels for the passage of ions. Due to their selective nature, such membranes are called differentially permeable or selectively permeable. Examples: Fish and other animals’ bladder, egg membranes, and plasma membrane of the cells.
{kind=link}
Imbibition
Imbibition is the phenomenon of adsorption of water or any other liquid, by the solid particles of a substance, without forming a solution. Solid substances or adsorbents (e.g., proteins and cellulose) which take part in imbibition are called imbibants. The liquid, usually water, which is imbibed is known as imbibate.
➢ Examples of Imbibition:
Jamming of wooden doors, windows, and drawers of desks during the rainy season is due to the imbibition of water by cellulose. Swelling of seeds, when immersed in water, is due to the imbibition of water by proteins.
➢ Characteristics of Imbibition
Amount of water imbibed by a substance is determined by the
● Water potential gradient between adsorbent and water.
● affinity of adsorbent for water.
Dry seeds have a highly negative water potential. On placing these seeds in water (Ψw = 0), a steep water potential gradient is set up. This results in rapid movement of water to the surface of dry seeds. This imbibition of water continues until an equilibrium is established between the water outside and in the seeds.
The process of imbibition has three important aspects:
01. Change in Volume: Due to imbibition, volume of imbibant increases, but total volume of water imbibed plus the imbibing material is less after imbibition due to compression of water.
02. Production of Heat: During imbibition, the water molecules get tightly adsorbed, and become immobilized. They lose most of their kinetic energy in the form of heat. It is called heat of wetting or heat of hydration.
03. Imbibition Pressure: When dry plant material, wood, or seeds are placed in water, they adsorb water and swell. The imbibed water produces a large pressure. This is called imbibition pressure. Now, this term has been replaced by matric potential which means water potential of the matrix. Air dried pea seeds, on coming in contact with water, can develop an imbibition pressure of upto 1000 bars (100 MPa).
Imbibition pressure is tested by placing dry seeds in a tin, adding water upto their upper level, and then tightly putting the lid over it. Within an hour, the lid will be blown off. alternately, fill a gas jar 3/4 with dry seeds. Pour water upto the brim. Cover the cylinder with a lid, and place a weight over the lid. Within a couple of hours the lid, along with weight, is lifted.
Proteins are powerful imbibants as compared to polysaccharides like starch, cellulose, or lignin. Pure gelatin adsorbs water more quickly than cellulose or lignin.
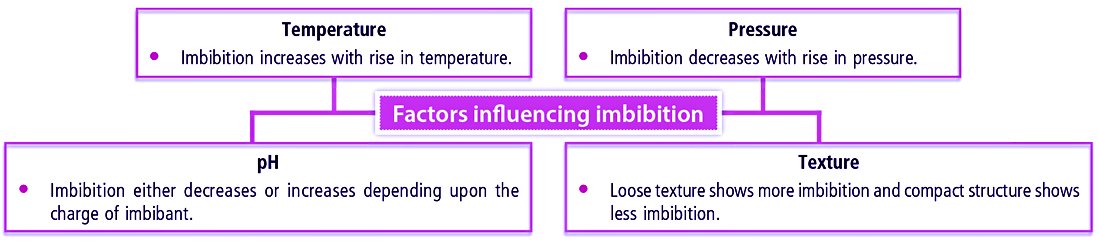
➢ Factors Affecting Imbibition
Factors influencing imbibition
● Looseness of the texture of imbibant shows more imbibition, while compactness of texture shows less imbibition.
● Imbibition rises with rising temperature.
● It decreases with rising pressure.
● Imbibition either decreases or increases, depending upon the charge of imbibant.
{kind=link}
➢ Importance of Imbibition to Plants
01. When dry seeds are soaked in water, they imbibe water and swell. This triggers the growth of the embryo. Thus, imbibition is the initial step in seed germination.
02. The seeds when soaked in water, imbibe water and swell up. Swelling results in the rupture of testa.
03. During absorption of water by roots, water is first adsorbed by the wall of root hairs and then by root hair.
04. Water moves by imbibition into the ripening ovule to form seeds.
05. Imbibition results in huge pressure developed by the swollen seeds. This pressure makes the seedlings emerge above the ground through the soil surface. This is called the imbibition pressure or matric potential.
In olden times, the imbibition pressure was used in breaking rocks and stones. Dry wooden stakes were driven into a crack in a rock and then soaked which develop enough pressure to split the rock.
Water Potential (Ψw)
Definition: Water potential is the difference in the free energy (chemical potential) of water molecules in the solution and that of pure water at the same temperature and pressure.
Water potential is denoted by the Greek symbol Ψ (Psi) or Ψw. It is measured in pressure units, such as bars, atmospheres, pascals (Pa) or megapascals (MPa) as in the case of osmotic pressure. The term ‘water potentiaľ was coined by Slatyer and Taylor (1960) in order to replace diffusion pressure deficit. [one MPa is equal to 106 pascals. A pascal (Pa) is a unit of measurement commonly applied to pressures—force per unit area].
Water always moves from the area of high water potential or high energy to the area of low water potential or low energy. The movement of water molecules continues until the water potential (Ψw) of two regions or two solutions becomes equal. The water potential of a solution can be determined using pure water as the standard of reference.
Pure water, at normal temperature and pressure, has a water potential of zero (0). The presence of solute particles reduces the free energy of water, and thus decreases the water potential (negative value). Therefore, the water potential of a solution is always less than zero. Hence, water always moves from the less negative potential (
➢ Components of Water Potential
Water potential (Ψw) in plant cells is determined by three internal factors: Matric potential (Ψm), Solute potential or osmotic potential ((Ψs), Pressure potential ((Ψp). A fourth factor, gravity potential (Ψg) also sometimes influences the water potential of cell sap, when the contents face tension, due to the force of gravity. The water potential in a plant cell or tissue can be written as the sum of solute potential, pressure potential and matric potential. Gravity potential (Ψg) is omitted as it is negligible as compared to the osmotic Ψs and Ψp .
Water potential (Ψw) = Solute potential (Ψs) + Pressure potential (Ψp) + Matric potential (Ψm)
01. Matric Potential (Ψm) : The colloidal particles present in the cell cytoplasm are hydrophilic in nature. These are macromolecules of carbohydrates, proteins and mucopolysaccharides. Water molecules are adsorbed to the surface of these macromolecules. This causes a decrease in the chemical potential of water because of immobilization. Matric potential is always a negative value and is measured in atmospheres.
The value of Ψw is maximum in young cells, fruits, seeds, and in plant cells living in extreme cold or dry conditions. But negligible in hydrated or mature cells. It is not significant in osmosis, so often disregarded. Thus, the equation may be simplified as
Ψw = Ψs + Ψp
2. Osmotic Potential or Solute Potential (Ψs) : It is defined, as the decrease in the water potential of a solution over its pure state, as a result of the presence of solutes. It is represented by SP or Ψs. Solute potential or osmotic potential (Ψs) is always in negative value. The more number of solute molecules in a solution is lower the solute potential (Ψs).
For a solution at atmospheric pressure, water potential (Ψw) will be equal to solute potential (Ψs).
Ψw = Ψs (at atmospheric pressure)
The non-electrolytic solutes (like carbohydrates, proteins, etc.) affect the chemical potential less than the electrolytic solutes. Ψs or SP is calculated by the following formula:
Ψs = C × R × T
where C = Concentration of solute particles in moles per liter of solution
R = Gas constant (it is equal to 0.083 bar or 0.082 atm)
T = Temperature (in Kelvin scale or absolute degrees)
It always has a negative value
3. Hydrostatic Pressure or Pressure Potential (Ψp) or Turgor Pressure (TP): Turgor pressure is the hydrostatic pressure with which the cell cytoplasm presses against the cell wall. It is also termed as hydrostatic pressure potential (Ψp).
Plant cell wall is elastic and exert pressure on the cell cytoplasm. This inward pressure of the cell wall is called wall pressure. The water molecules that diffuse into the cells enter the vacuole and produce hydrostatic pressure or turgor pressure (TP) in the cell sap.
Due to turgor pressure, the protoplast of a plant cell will press the cell wall to the outside. The cell wall, being elastic, presses the protoplast with an equal and opposite force. The force exerted by the cell wall over the protoplast is called wall pressure (WP).
With the continuous entry of water into the cell, the wall pressure keeps increasing until the cell becomes turgid. In a fully turgid cell, wall pressure (WP) is equal and opposite to turgor pressure (TP) and prevents further entry of water, i.e., WP = TP. In this state, the value of wall or turgor pressure becomes equal to osmotic potential, Ψp = Ψs. The state of plant cells being turgid or swollen due to endosmosis is called turgidity.
The basic driving force in osmosis is the difference in the free energy of water on two sides of semipermeable membrane. An increase in the temperature or pressure increases the free energy and pressure potential of the solution.
An increase in the solute concentration lowers the free energy and decreases the pressure potential of the solution.
{kind=link}
► Significance of Turgor Pressure (Turgidity)
01. Turgor pressure is essential for plants to live and grow.
02. Turgor pressure helps in the opening and closing of stomata by regulating the turgidity of the guard cells.
03. Turgor pressure keeps cell organelles stretched and expanded. It keeps leaves, flowers, fruits, etc., expanded.
04. Turgor pressure provides mechanical support to non-woody tissues (e.g., parenchyma).
05. Loss of turgidity leads to wilting of leaves and drooping of shoots.
06. Turgor pressure helps in the elongation and enlargement of cells during growth and it keeps leaves erect and fully expanded, and properly oriented to light.
07. Leaf movements of Mimosa pudica (the touch-me-not plant) are due to turgidity.
Osmosis
Osmosis is a special type of diffusion of water that occurs through a semipermeable membrane. Osmosis is movement of solvent or water molecules from the region of their higher diffusion pressure or free energy to the region of their lower diffusion pressure or free energy, across a semipermeable membrane. The phenomenon of osmosis was discovered by Nollet in 1748.
Following definitions have been given for osmosis:
01. Osmosis is the diffusion of water from its pure state or from a dilute solution to a solution of higher concentration when the two are separated by a semipermeable membrane.
02. Osmosis is the movement of water from its higher chemical potential (i.e., from pure state or dilute solution) to its lower chemical potential (in a strong solution) through a semipermeable membrane.
03. Osmosis is the movement of water or solvent molecules from the region of their higher diffusion pressure (or free energy) to the region of lower diffusion pressure (or free energy) across a semipermeable membrane.
The direction and rate of osmosis depend upon the sum of the pressure gradient (gradient of Ψp) and concentration gradient (gradient of Ψs). The gradient is determined by the difference in water potential of solutions separated by a semipermeable membrane. A solution in which water enters osmotically is called an osmotically active solution. This solution has a lower water potential. Diffusion of water into it will continue through the membrane till an equilibrium is reached. When the equilibrium reaches, water potential becomes equal on both sides of the membrane.
► Requirements of Osmosis
01. Semipermeable Membrane: It is essential for the operation of osmosis because it allows the passage of water without allowing the movement of solutes through it. In plants, biomembranes function as semipermeable membranes.
02. Concentration of Dissolved Solutes: The movement of water occurs in osmosis from higher solute potential, i.e., less negative solute potential to more negative solute potential.
03. Pressure Difference: Pressure difference affects solute potential. Thus, a positive pressure applied over an osmotically active solution reduces the entry of water into it, whereas a similar pressure applied over an external solution or water may enhance the passage of water.
► Factors influencing Osmosis
● Presence of imperfect semipermeable membrane disturbs osmosis.
● The concentration of dissolved solute on both sides of the semipermeable membrane also influences osmosis.
● Osmosis is also influenced by differences in pressure.
► Mechanism of Osmosis
When solute particles are added to water, they decrease its chemical potential by decreasing the mole fraction of water. In osmosis, a semipermeable membrane separates dilute solution A and concentrated solution B, but the solute particles cannot pass through it. Water molecules being in random motion, strike semipermeable membrane on both sides and pass through the same. Since more free water molecules are present on the side of dilute solution A, more molecules pass through the membrane into solution B as compared to the reverse flow. Thus, there is a net diffusion of water from its higher chemical potential (dilute solution) to its lower chemical potential (concentrated solution).
{kind=link}
► Significance of Osmosis
01. It helps in absorption of water by root hairs.
02. It leads to cell-to-cell movement of water.
03. It induces turgidity in cells which helps in stretching of stem and in keeping the leaves expanded.
04. Osmosis provides mechanical support in nonwoody plants like maize, sugarcane and banana by making the cells fully turgid.
05. Growing tips of roots remain turgid because of osmosis and thus are able to penetrate into the soil.
06. Osmosis regulates the opening and closing of stomata.
07. Resistance of plants to drought and frost is brought about by osmosis.
08. Movement of plant and plant parts is brought about by cell’s turgor due to osmosis, for example, movement of leaflets of the sensitive plant, Mimosa pudica.
09. High osmotic pressure protects the plants against drought and frost injury.
10. Seeds and spores are able to pass through unfavourable periods due to high osmotic pressure.
► Types of Osmosis
{kind=link}
Osmosis is of two types – endosmosis and exosmosis.
01. Endosmosis: The osmotic entry of water into a cell, organ or system is called endosmosis. It happens when a cell or osmotic system is placed in pure water or hypotonic solution. Endosmosis creates a turgor or hydrostatic pressure. An increase in volume of the cell or osmotic system also takes place. It makes the cell turgid.
Example: Put some raisins in a dish full of water. Observe after a few hours. These have swollen due to the osmosis of water. In raisins, there is a stronger solution of sugar than that of the outer solution of water and the outer membrane of raisins serves as a semipermeable membrane. Thus, water enters into the raisins and makes them turgid. This shows endosmosis.
02. Exosmosis: The osmotic withdrawal of water from a cell, organ, or system is called exosmosis. It decreases the volume of cell or of the osmotic system. Because of exosmosis, the cell becomes deplasmolysed and flaccid.
Example: Place some swollen raisins or fresh grapes with intact stalks in 10% sugar solution in a petri dish. Grapes (raisins) shrink in size and become wrinkled. The loss of water in response to the external hypertonic solution is called exosmosis.
► Osmotic Concentration of Solutions
{kind=link}
A solution that can cause an osmotic entry of water into it is called an osmotically active solution.
Depending upon the osmotic concentration, the solution can be of three types:
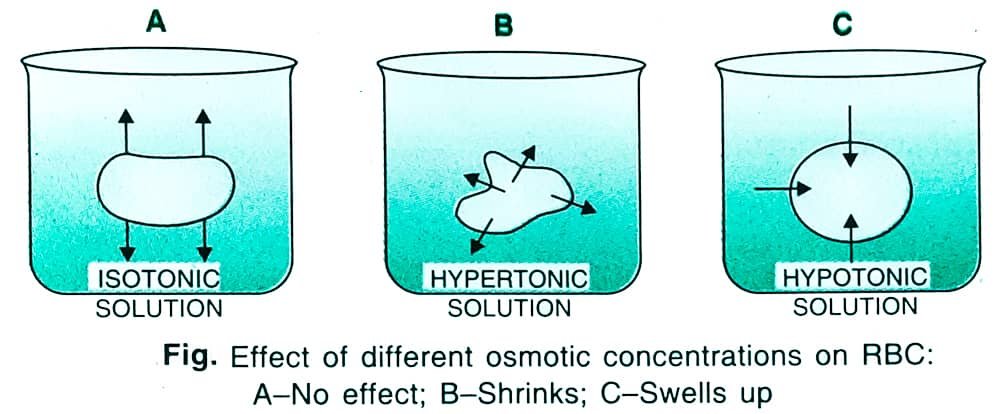
01. Isotonic Solution: The two solutions with the same osmotic concentration are named as isotonic solutions. When separated by a semipermeable membrane, an isotonic solution neither gains nor loses water. Cells placed in isotonic solution also do not undergo any change. They remain flaccid.
02. Hypertonic Solution: A solution having high osmotic concentration as compared to another solution is known as a hypertonic solution. It withdraws water from the other solution when the two are separated by a semipermeable membrane. A cell placed in hypertonic solution will shrink and undergo plasmolysis due to the loss of water from it.
03. Hypotonic Solution: A solution having a low osmotic concentration as compared to another solution is known as a hypotonic solution. When separated by a semipermeable membrane, the hypotonic solution will allow the passage of water from it to another solution. A cell placed in a hypotonic solution will swell up and become turgid.
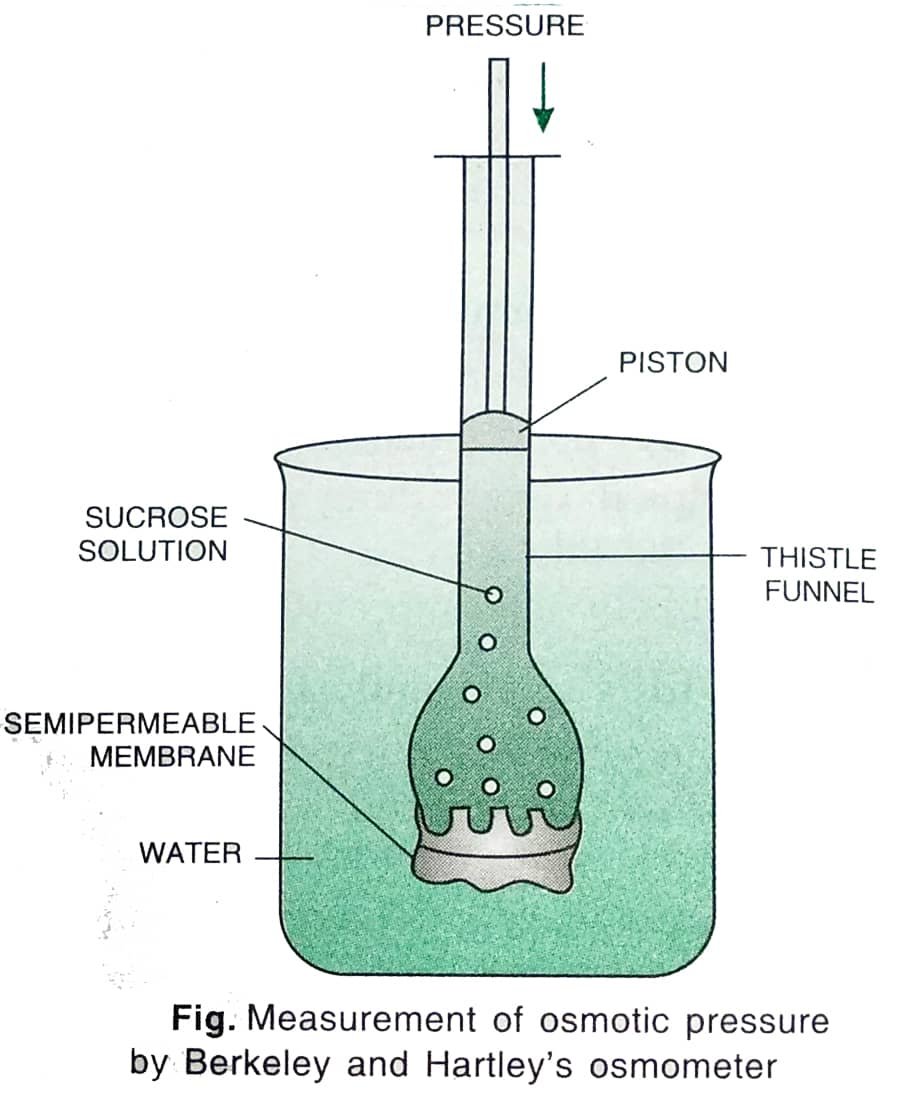
— When a solution is separated from pure solvent (water) by a semipermeable membrane, the Solvent molecules begin to move towards the solution due to osmosis. This creates a hydrostatic pressure in the solution which is called osmotic pressure. Osmotic pressure is the maximum pressure developed in an osmotically active solution when it is separated from pure water by a semipermeable membrane under ideal conditions of osmosis. — Osmotic pressure of a solution is equal to the external pressure required to stop the entry of water into the solution. It is represented by the symbol 𝞹 (pie). It is measured in atmosphere (atm) or bars or pascals (Pa) and megapascals (MPa) [One bar = 0.987 atm = 10 pascals (Pa) = 0.1 megapascal (MPa)]. Osmotic pressure is numerically equal to osmotic potential or solute potential (Ψs). The osmotic pressure has a positive value while osmotic potential has a negative value, i.e., Ψs = - 𝞹 ■ Measurement of Osmotic Pressure Osmotic pressure is measured by an osmometer. Commonly used osmometers are Berkeley and Hartley's osmometer, and Pfeffer's osmometer. In the former osmometer, pressure is applied to prevent the entry of water in the solution of the thistle funnel separated from the semipermeable membrane. ■ Osmotic Pressure (OP) in Some Plants 1. OP in halophytes = 202 atm 2. (a) OP in xerophytes = 10–30 atm (b) OP in xerophytes under draught conditions = up to 60 atm 3. OP in mesophytes = 5–15 atm 4. OP in hydrophytes = 1–3 atm Thus, the osmotic pressure of halophytes is the highest and of hydrophytes is the lowest. ■ Reverse Osmosis When the pressure applied to the solution is more than the osmotic pressure, water starts to flow out of the solution into the surrounding water. This movement of water is against the concentration gradient and is called reverse osmosis. This process is used to de-saline water by removing salt from sea water for use in chemical industries and for drinking water.
{kind=link}
Plasmolysis
Shrinkage of the protoplast of a cell, from its cell wall, under the influence of a hypertonic solution is called plasmolysis. Due to the withdrawal of water from the cytoplasm and central vacuole of the cell, the size of the protoplast reduces. This is the first stage of plasmolysis called, limiting plasmolysis. The hypertonic external solution continues to withdraw water from the central vacuole and protoplast, by exosmosis, which withdraws itself from the corners and results in incipient plasmolysis. At this stage, the hypertonic solution enters the cell in between the protoplast and the cell wall. Due to continued exosmosis, protoplast shrinks further and withdraws from the cell wall except at very few points. It is known as evident plasmolysis.
► Deplasmolysis
If a plasmolysed cell is again placed in hypotonic solution or a solution of less concentration than the cell sap, water from outside enters the protoplasm or endosmosis occurs. As a result the protoplasm again comes back to its original position and stretches up with the cell wall. This is termed deplasmolysis. If a cell is placed is an isotonic solution, i.e., a solution whose concentration is just equal to the concentration of the cell sap, neither shrinking nor swelling of the protoplasm will occur. Plasmolysis is a kind of defense mechanism found in plants to protect themselves against adverse conditions.
{kind=link}
01. It is a vital phenomenon as it explains the process of osmosis.
02. It proves the permeability of the cell wall and the semipermeable nature of the protoplasm.
03. It helps in determining whether a particular cell is living or dead since the plasmolysis does not take place in a dead or nonliving cell.
04. It helps in determining the osmotic pressure of a cell.
► Practical Utility of Plasmolysis
01. It is involved in the preservation of meat, jellies, and other foodstuffs. Salting of the foodstuffs (to be preserved) raises the osmotic pressure of the preservating medium and bacteria and fungi get plasmolysed and killed due to exosmosis.
2. Plasmolysis is also involved in the killing of weeds.
3. The phenomenon is useful in preventing the growth of plants in the cracks of walls.
The difference in the concentration of solutions, on two sides of a semipermeable membrane, results in the flow of water from a solution of low concentration to a solution of higher concentration. When a cell is placed in a hypotonic solution, water enters into the cell and the turgor pressure of the cell increases. As a result of the increase in TP, the wall pressure of the cell also increases (equal in magnitude but opposite in direction). (i) In fully turgid cells, turgor pressure is equal to osmotic pressure, i.e., TP = OP. Therefore DPD is zero. When the cell is fully turgid, DPD = OP - TP DPD = OP - OP (as TP =OР) DPD = 0 (ii) In a flaccid cell, turgor pressure is zero and wall pressure is negative. Therefore, DPD is equal to osmotic pressure and more than wall pressure. (a) When TP = 0, DPD = OP - TP DPD = OP - 0 DPD = OP (b) When wall pressure is negative, DPD = OP - (-WP) DPD = OP + WP (iii) When a flaccid cell is placed in pure water, the water moves into the cell causing turgor pressure (TP) to develop and a decrease in the osmotic pressure of cell sap. During this condition, DPD becomes equal to OP – TP, i.e., DPD = OP - TP
[/su_note]
OSMOTIC RELATIONS OF PLANT CELLS
A plant cell functions as an osmotic system. It lies in a water medium or lies in contact with other cells having water. The vacuole is filled with cell sap. The cell sap is an aqueous solution of salt, sugar, and organic acids in a fairly high concentration. It is osmotically active. It can gain or lose water depending upon the diffusion pressure of the surrounding medium. The solution of the surrounding medium can be hypotonic, hypertonic or isotonic to the cell sap.
The osmotic relationship of a plant cell with its surroundings is represented by cell turgidity.
{kind=link}
01. Rise in Turgidity (Gain of Water)
When a flaccid cell is placed in a hypotonic solution, water starts entering the cell by osmosis. Continuous entry of water results in hydrostatic pressure called the turgor pressure (TP) in the cell. This forces the protoplasm to press the cell wall towards the outside. Turgor pressure continues to rise as more water flows in. In the end, a stage comes when the cell becomes fully turgid or distended.
The force with which the water is drawn into the cell is called diffusion pressure deficit (DPD) or water potential (Ψp). In a fully turgid cell, the turgor pressure becomes equal to osmotic pressure and DPD becomes zero. Now, no entry of water takes place.
Ψw = Ψs + Ψp = 0
02. Loss of Turgidity (Loss of water)
When a plant cell is placed in a hypertonic solution, water moves out of the cell because of the higher chemical potential of the cell sap. The vacuole shrinks in size and protoplasm is no longer pressed against the cell wall. Now, both the turgor pressure and wall pressure become zero. Such a cell is called plasmolysed and the phenomenon as plasmolysis. Such a cell becomes flaccid and TP of such a cell becomes zero. If the external solution does not cause any further exosmosis, the value of its solution potential (Ψs) will be equal to or isotonic to the solute potential of the cell.
Ψw = Ψs + Ψp or Ψw = Ψs
03. Water Movement Between Adjacent Cells
The direction and rate of water movement between two adjacent cells depend upon their water potential or DPD. The direction of water flow is always from the cell with lower DPD (or higher water potential) to the cell with higher DPD (or lower water potential), even though the osmotic pressure of the former may be higher than that of the latter.
[/su_note]
WATER ABSORPTION
01. SOIL WATER
Plants absorb water from the soil with the help of their roots. This water is found in different forms in the soil. The soil plays important role in water storage and serves as a medium for root growth. It is also a reservoir of mineral nutrients and provides anchorage for plants. The ultimate source of water for the soil is rain or irrigation. A part of rainwater does not enter the soil, but is drained away from the soil surface, along the slope and is called run-away water or run-off.
The optimum amount of water that a soil can hold without disturbing its aeration is called its field capacity. Water in the soil is known as soil water or soil solution. The value of field capacity is 25-35% in loam soil. The total water present in the soil is called holard. Water available to plants for absorption is called chresard (available water). Soil water that cannot be absorbed by plants is called echard (unavailable water).
If soil contains only unavailable water, the plants growing in it will undergo permanent wilting. It is a stage of loss of turgidity when plants do not recover even in saturated conditions unless they are supplied immediately with water.
■ Permanent Wilting Percentage
It is a stage of loss of turgidity when plants do not recover even in saturated conditions and become permanently wilted and die. The percentage of water present in the soil at the time of permanent wilting is called permanent wilting coefficient. At permanent wilting percentage, the soil contains about 10% of water which is either present in fine micropores or in the nonavailable stage. Water present in the soil in excess of its field capacity causes waterlogging. The growth of plants is poor in waterlogged soils due to reduction in aeration, inactivation of aquaporins, and poor growth of root hairs.
■ Types of Soil Water
The water present in the soil is in the following states:
{kind=link}
01. Capillary Water: This is the only water available to plants. It is held between soil particles (diameter ≤ 20 mm) by capillary force. It contains a number of mineral salts in solution.
02. Hygroscopic Water: The soil particles are surrounded by a film of water called hygroscopic water. This water is not available to the plant as it is not free but remains around colloidal soil particles due to hygroscopic action.
03. Chemically Combined Water: A small amount of water is chemically bound with soil particles (e.g., hydrated oxides of aluminium, iron, silicon, etc.) is said to be the combined water. This water is not available to plants.
04. Gravitational Water / Free Water: When water enters the soil from the surface either by irrigation or rain, it percolates downward through the spaces between the soil particles under the influence of gravitational force and reaches the water table. This is called gravitational water. It is not available to plants because it lies far below the reach of the roots.
Soil stratum containing groundwater is called aquifer. The upper layer of aquifer or groundwater is called water table. Wells, tube-wells, handpumps, etc., draw water from this layer.
05. Run off Water / Run away Water: After heavy rainfall, all the water does not enter the soil and gets drained away along the slopes. This is called run off water and is not available to the soil or plants.
02. LONG DISTANCE MOVEMENT OF WATER
The path of water movement in plants is through the vascular bundles, especially through the xylem.
Both, the substances and water have to travel a long distance in large trees. This long distance transport cannot occur by diffusion alone because diffusion is a slow process. For example, the movement of a molecule across a typical plant cell, i.e., about 50 μ, takes approximately 2.5 seconds. At this rate, the movement of molecules over a distance of one meter may take several years. Thus, plants have developed a mass system or bulk-flow system. It is the movement of substances in bulk from one region to another due to the pressure difference between the source and sink. In bulk or mass flow, all the dissolved or suspended substances travel at the same pace in the flowing water of a river.
The long distance mass or bulk movement of substances that occur through vascular tissues of plants is called translocation. Xylem and phloem are the conducting or vascular tissues of plants. Xylem translocates mainly water and mineral salts from roots to the aerial parts of the plant. Phloem translocates mainly organic solutes from leaves to other parts of the plant. Bulk flow is achieved either through a positive hydrostatic pressure gradient in phloem as in a garden hose pipe or a negative hydrostatic pressure gradient in xylem like suction through a straw.
TABLE Movement of Substances Through the Plant
| Substances | Uptake | Movement | Eliminations | |||
| Root | Mechanism | Root | Mechanism | Shoot | Mechanism | |
| 1. Water | Root hair | Osmosis | Xylem | Mass flow | Stomata, cuticle and lenticels | Diffusion of water vapour |
| 2. Solutes | Root hair, leaves (photosynthesis) | Diffusion or active transport | Xylem (inorganic), phloem (organic) | Mass flow | Shedding of leaves, bark, fruits, and seeds or retained till death | Controlled by plant growth substances |
| 3. Gases | Stomata, lenticels and epidermis | Diffusion | Intercellular spaces and through cells | Diffusion | Stomata, lenticels, roots, epidermis | Diffusion |
03. WATER ABSORPTION SYSTEM
Land plants absorb water through the root system. A deep root system occurs in trees and plants of dry areas where the upper soil layer is nearly dry. Some desert plants like Prosopis, Tamarix, etc., also have long roots to reach the water table. They are called phreatophytes.
In herbaceous plants, the root system is spread out in upper layers of soil and is highly branched. The root hair zone is the zone of rapid water absorption. It lies some 20 – 200 mm above the root tip and bears thousands of root hairs.
Root hairs are unicellular tubular extensions from epiblema cells. They are about 50-1500 μm (i.e., 0.05-1.5 mm) in length and about 10 μm in width. Each root hair has a central vacuole filled with osmotically active cell sap and a peripheral cytoplasm. The wall is thin and permeable with pectic substances in the outer layer and cellulose in the inner layer. The root hairs passing into capillary micropores between soil particles absorb capillary water.
■ Mycorrhizal Water Absorption: In mycorrhiza, a large number of fungal hyphae are associated with the young roots. They form a network around the young root or penetrate the root cells. They provide minerals and water to the roots and roots provide sugars and nitrogen containing compounds to the mycorrhizae. Some plants have an obligate association with mycorrhizae. For example, Pinus and orchid seeds do not germinate without mycorrhizal association.
04. MECHANISM OF WATER ABSORPTION
Water molecules are first imbibed by the pectic layer of the cell wall of root hairs and then absorbed either by passive absorption or active absorption.
The process of active and passive absorption of water by root was described by Kramer (1937).
(i). Passive Water Absorption
In passive absorption of water, root cells play a passive role, because the forces responsible for water absorption by roots originate in the leaves due to transpiration. Rapid evaporation of water from the leaf surface reduces the turgor pressure of mesophyll cells resulting in an increase in their diffusion pressure deficit (DPD). Mesophyll cells then withdraw water from inner mesophyll cells, which draw water from the xylem. This causes the development of suction pressure and pulls water inside xylem vessels. It is called transpiration pull.
Since there is a continuous column of water from leaves to roots and to root hair through cells of endodermis and cortex, the DPD is transferred to root hair. Due to the difference in water potential between soil water and xylem vessels, water moves across by the suction force as a continuous column.
Passive absorption has been further supported by the following points:
➢ It is a regulated process and the rate of absorption is under the control of transpiration.
➢ It satisfies critically the movement of water against osmotic gradience.
➢ The rate of absorption is almost equal to the rate of transpiration.
➢ In rapidly transpiring plants, the cut ends of the stem are capable of absorbing water.
➢ It is a purely physical process.
➢ The movement of water across cells can be much faster as against simple osmotic diffusion. Thus, it satisfies as to how the excess demand of water is met during fast transpiration
(ii). Active Water Absorption
Active absorption of water takes place due to the activities in the root and is not concerned with the force generated in the shoot. It requires an expenditure of metabolic energy i.e. ATP. Two mechanisms have been given to explain active water absorption:
(a) Osmotic Mechanism of Active Water Absorption
The osmotic pressure or osmotic potential of soil water plays an important role in the absorption of water. The osmotic pressure of soil water remains below 1 atm, but that of cell sap is 2–8 atm. Due to the water potential gradient between the soil solution and the cell sap, absorption of water does not require any expenditure of energy. Water enters into the cell sap of root hair cells and from there into other epidermal cells till the water potential of the root cell sap is more negative than that of the soil solution. Water absorbed by root hairs moves from cell to cell along the concentration gradient (or water potential gradient) and reaches the endodermis and pericycle. From here, water enters the xylem elements and is translocated upwards.
(b) Nonosmotie Mechanism of Active Water Absorption
This type of water absorption by root hairs takes place at the expense of energy. It occurs against diffusion pressure gradient. Energy is needed for the absorption and accumulation of salts in the xylem and increasing the permeability of cells.
The following points have been placed in support of this theory:
(i) Wilting of roots occurs in nonaerated soils such as flooded areas.
(ii) Respiratory inhibitors such as KCN reduce the rate of water absorption and exudation from the cut end of stems. This shows some correlation between the two processes.
(iii) Occurrence of day and night variations in water uptake and root pressure.
(iv) Low auxin concentration increases water intake and exudation.
(v) Process of water absorption occurs in living cells.
Root pressure is the manifestation of active water absorption.
TABLE Differences between Passive and Active Water Absorption
Sl No Active Absorption Passive Absorption 01 The active absorption of water takes place due to the activity of root and root hairs. It occurs due to the activity of the upper part of the plant, such as shoot and leaves. 02 The absorption of water occurs by the osmotic and non-osmotic processes. The water is absorbed due to the active transpiration in aerial parts. 03 The root hairs have high DPD as compared to soil solution and therefore, water is taken in. The absorption occurs due to tension created in xylem sap by transpiration pull. Thus, water is sucked in by the tension. 04 In the movement of water, living part of protoplasts (symplast) is involved. The movement of water is through free spaces or apoplast of root and it may include cell wall and intercellular spaces. 05 The rate of absorption depends upon DPD or the difference in osmotic concentrations between the two. The rate of absorption depends upon transpiration. 06 It uses metabolic energy, i.e., ATP. It uses solar energy. 07 It does not occur in the absence of roots. It can occur even in the absence of roots. 08 Living root cells pump water into the xylem. Thus, it is manifested in the form of root pressure. Tension is built up in the xylem due to transpiration. It is manifested in the form of a transpiration pull. 09 The movement of water is against the concentration gradient. The movement of water is in accordance to osmotic or
concentration gradient. 10 It usually occurs at night, when transpiration stops, due to the closure of stomata. It occurs during the day, when transpiration is going
on, and stops at night, when transpiration stops. 11 Bleeding and guttation are closely associated with active absorption. Transpiration is associated with passive absorption. 12 It is influenced by inhibitors (metabolic poisons). It is not influenced by inhibitors (metabolic poisons).
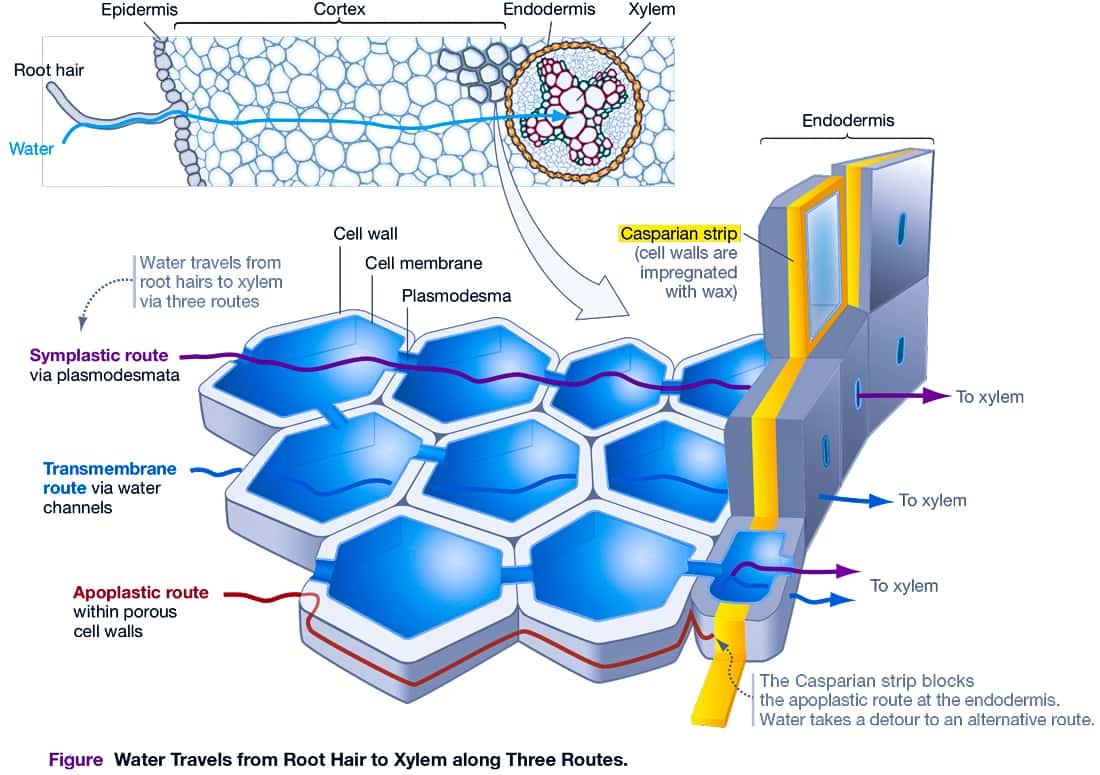
05. PATHWAYS OF WATER MOVEMENT IN ROOTS
To understand how water enters a root, consider the cross section through a young buttercup root. Starting at the outside of the root and working inward, notice that several distinct tissues are present:
● The epidermis (literally, “outside skin”) is a single layer of cells. In addition to protecting the root, some epidermal cells
produce root hairs, which greatly increase the total surface area of the root.
● The cortex consists of ground tissue and stores carbohydrates.
● The endodermis (“inside skin”) is a cylindrical layer of cells that forms a boundary between the cortex and the vascular tissue. The function of the endodermis is to control ion uptake and prevent ion leakage from the vascular tissue.
● The pericycle (“around circle”) is a layer of cells located just interior to the endodermis that forms the outer boundary of
the vascular tissue. The pericycle can produce lateral roots.
● Conducting cells of the vascular tissue transport water and nutrients between roots and shoots and are located in the
center of roots in buttercups and other eudicots. Notice that, in these plants, phloem is situated between each of four arms formed by xylem, which is arranged in a cross-shaped pattern.
■ Three Routes through Root Cortex to Xylem
When water enters a root along a water-potential gradient, it does so through root hairs. As water is absorbed, it moves through the root cortex toward the xylem along three distinct routes.
{kind=link}
(i). Apoplastic Movement / Apoplast Pathway
The apoplastic pathway is outside the plasma membranes. The apoplast consists of cell walls, which are porous, and the spaces that exist between cells. Water moving along the apoplastic route must eventually pass through the cytoplasm of endodermal cells before entering the xylem.
In this pathway, water passes from root hairs to xylem through nonliving cell walls without crossing the living cell membranes or cytoplasm.Major portion of water in the root cortex moves by apoplast because this pathway provides the least resistance to the movement of water. However, beyond the cortex, apoplast movement of water is blocked by casparian strips made of impermeable lignosuberin in the walls of endodermal cells.
(ii). Symplastic Movement / Symplast Pathway
The symplastic pathway is inside the plasma membranes. The symplast consists of the cytosol and the continuous connections through cells via plasmodesmata.
In this pathway, water passes from cell to cell through cytoplasm via cell membrane and plasmodesmata without entering cell vacuoles. Beyond endodermis, water moves through the cell membranes and plasmodesmata. It is also called the transmembrane pathway.
Symplastic movement may be aided by cytoplasmic streaming. Cytoplasmic streaming movements can be observed in the cells of Hydrilla leaf by the streaming movement of chloroplasts. Symplastic movement is, however, slower than apoplastic movements.
(iii). Transmembrane Pathway
The transmembrane pathway is based on flow through aquaporin proteins—water channels located in the plasma membranes of many cells. Some water may also diffuse directly across plasma membranes.
In essence, water can flow through tissues by passing through the symplast via plasmodesmata, crossing membranes via aquaporins, or passing around cells via the apoplast.
■ The Role of the Casparian Strip
The situation changes when water reaches the endodermis. Endodermal cells are tightly packed and secrete a narrow band of wax called the Casparian strip. This layer is composed primarily of a compound called suberin, which forms a waterproof barrier where endodermal cells contact each other.
The Casparian strip blocks the apoplastic pathway by preventing water from moving through the walls of endodermal cells and into the vascular tissue. The Casparian strip does not affect water and ions that move through the symplastic route.
The Casparian strip is important because it means that for water and solutes to reach vascular tissue, they have to move
through the cytoplasm of an endodermal cell. Endodermal cells, in turn, act as gatekeepers by regulating what enters the xylem.
By forcing water and ions to cross at least two membranes on their way from the soil to the xylem, one entering the symplast outside the Casparian strip and one leaving the symplast inside the Casparian strip, plants can use a specific channel and carrier proteins to control what moves to the shoots. Endodermal cells allow ions such as potassium (K+) that are needed by the plant to pass through to the vascular tissue. In contrast, these cells can limit the passage of ions such as sodium (Na+) or heavy metals that are not needed or may be harmful.
TABLE: Differences between Apoplast and Symplast Pathways of Water Movement
| Sl No | Apoplast Pathway | Symplast Pathway |
| 01 | It consists of nonliving parts, i.e., cell walls and intercellular spaces. | It consists of cytoplasm and plasmodesmata. |
| 02 | Water does not enter the living cells. | Water enters the living cells either in the area of root hair cells or endodermis. |
| 03 | There is little resistance in the movement of water. | There is some resistance in the movement of water through the symplast. |
| 04 | It is a faster process. | It is relatively a slower process. |
| 05 | It is not affected by the metabolic activity of the root. | It is influenced by the metabolic status of the root. |
■ Factors affecting Water Absorption
► EXTERNAL FACTORS
(i) Available soil water – It is optimum at field capacity, water absorption decreases above it. It begins to decline and stop at PWP.
(ii) DPD of soil water – Normal DPD of soil water is 0.1 -0.3 atm. Above 0.1 atm the rate of water absorption begins to decline. Near 4.0 atm, water absorption falls below requirement.
(iii) Soil air – Oxygen content of soil air is generally less than that of atmosphere while carbon dioxide content is higher. The rate of water absorption begins to decline below 15% oxygen and more than 1% carbon dioxide.
(iv) Transpiration – It creates a force for passive absorption of water. When transpiration is very high, the rate of water absorption declines due to the development of root-soil resistance and contraction of the root.
(v) Soil temperature – Optimum soil temperature for water absorption is 15-25°C. A soil temperature of 30-35°C decreases water absorption. A lower temperature similarly reduces water absorption.
Internal factors
► INTERNAL FACTORS
(i) Extent of root system – The amount of water absorption is directly dependent on the extensiveness of the root system especially the number of rootlets and their root hairs.
(ii) Growth and metabolism of root – Root hairs have a short life span. Continued water absorption depends upon growth of root branches and the formation of new root hairs.
(iii) Depth of root system – Deeper roots are less efficient than the upper ones.
(iv) Internal resistance – Water absorbed by root hairs travels to the root Interior and from there it rises upwardly. The rate of this transfer is inversely related to the degree of internal resistance.
● Atmometer is an apparatus consisting of an unglazed porous pot connected to a narrow glass tube. The apparatus is filled with air free water. The free end of the glass tube is dipped in beaker containing mercury. As water evaporates from the porous pet, mercury begins to rise upwards in the glass tube showing a pull due to evaporation. (Similar to pull by transpiration)
● Tensiometer is an instrument for measuring soil water tension.
● Psychrometer is an instrument for measuring both relative humidity and transpiration.
● Manometer is an instrument for measuring pressure or tension in gases or liquids.
● Barometer is used for measuring atmospheric pressure.
● Barograph is used for the recording of barometer,
[/su_note]
ASCENT OF SAP (TRANSLOCATION OF WATER)
Water absorbed by plants is actually a dilute solution of minerals and is called sap. The upward movement of water or sap (water with dissolved ingredients) from the root to the top of the plant is called ascent of sap. Roots are organs concerned with the absorption of water. If the plant is to survive and grow, this water absorbed by roots must reach the top of plants to replace the water lost in transpiration and to be used in photosynthesis as a raw material.
The upward movement of water occurs through the tracheary elements (tracheids and vessels) of xylem. Sap is lifted from near the root tip to the shoot tip against the force of gravity, sometimes to great heights. Some trees are much taller, often attaining a height upto 110-130 m, e.g., Sequoia sempervirens (height 110 m), Picea sitchensis (height 95-100 m), Pseudotsuga menziesii (height 100 m), Eucalyptus (height above 130 m), etc. The force required to move water to such heights are substantive. The rate of ascent of sap (= translocation) is 25–75 cm/min (15–45 m/hr).
Several theories have been put forward to explain the mechanism of the ascent of sap. The four main theories are:
● Vital force theory,
● Root pressure theory,
● Capillary force theory, and
● Cohesion-tension and transpiration pull theory
01. VITAL FORCE THEORY
A common vital force theory about the ascent of sap was put forward by Sir J.C. Bose in 1923. It is called Pulsation theory. According to this theory, the living cells of the innermost layer of the cortex just outside the endodermis absorb water from the outer side and pump it into xylem channels. But water continues to rise upwards in the plant in which roots have been cut or the living cells of stem are killed by poison or heat (Strasburger, 1891). Therefore, living cells do not seem to be involved in the ascent of sap.
02. ROOT PRESSURE THEORY
This theory was proposed by Priestley (1916). According to this theory, root pressure is a hydrostatic pressure that develops into the root xylem due to the active absorption of water. It is a positive pressure that is maintained by the activity of living cells.
The Casparian strip in endodermal cells is essential for root pressure to develop. Without an apoplastic barrier between the xylem and the environment, ions and water would simply leak out of roots.
Stomata normally close during the night, when photosynthesis is not occurring and CO2 is not needed. Their closure minimizes water loss and slows the movement of water through plants. But roots often continue to accumulate ions that their epidermal cells acquired from the soil as nutrients, and these nutrients are actively pumped into the xylem. The influx of ions lowers the water potential of xylem below the water potential in the surrounding cells. As a result, water from nearby cells enters xylem.
As water flows into xylem from other root cells in response to the solute gradient, a positive pressure is generated at night that forces fluid up the xylem. More water moves up xylem and into leaves than is being transpired from the leaves.
In certain low-growing plants, such as strawberries, enough water can move to force water droplets out of the leaves, a phenomenon known as guttation.
■ Objections to Root Pressure Theory:
At one time, positive root pressure formed the basis of a leading hypothesis to explain how water moves from roots to leaves in trees. However, research showed that over long distances, such as from the ground to the top of a large tree, the force of root pressure is not enough to overcome the force of gravity on the water inside xylem.
In addition, researchers demonstrated that cut stems, which have no contact with the root system, are still able to transport water to leaves. Biologists concluded that there must be some other mechanism involved in the long-distance transport of water.
Root pressure has been observed in herbaceous plants but it is not sufficient to drive water to a distance of 400 ft in the trunks of tall trees as is evidenced by the following findings:
(i). The magnitude of root pressure has been generally found to be under 2 atm while a pressure of about 20 atm is needed to raise water to the tops of tall trees.
(ii). The rate of sap exudation is not adequate as compared to the rate of passage of sap through the xylem. Root pressure is generally observed at night when transpiration is low.
(iii). Root pressure has been found to be the lowest during summer when the rate of transpiration is very rapid and the need for upward conduction of water is very high.
(iv). Water continues to rise even in the absence of root pressure.
(v). Root pressure is seen only during the most favourable periods of growth like spring or rainy season. At this time, cell sap is strongly hypertonic to soil solution and the transpiration rate is low. In summers, when water requirements are high, root pressure is generally absent.
(vi). Finally, the observation that xylem sap, under normal conditions is under tension instead of pressure proves that root pressure is not an important factor in the translocation of water.
03. CAPILLARY FORCE THEORY
Capillary force theory was given by Boehm (1809). According to this theory, water rises up in narrow tubes of xylem vessels by (i) adhesion, (ii) cohesion, and (iii) surface tension.
Capillary action results when adhesion creates an upward pull at the water–container interface, surface tension creates an upward pull all across the surface, and cohesion transmits both forces to the water below. All three forces counteract the effect of gravity, and the result is capillary action.
➢ Adhesion is a molecular attraction among unlike molecules.In this case, water interacts with a solid substrate—such as the glass walls of a capillary tube or the cell walls of tracheids or vessel elements—through hydrogen bonding. Water molecules are pulled upward as they bond to each other and adhere to the side of the tube.
➢ Cohesion is a molecular attraction among like molecules, such as the hydrogen bonding that occurs among molecules in water. Because water molecules cohere, the upward pull by adhesion is transmitted to the rest of the water
column. The water column rises against the pull of gravity.
The effects of adhesion, cohesion, and gravity are responsible for the formation of a concave surface boundary called a meniscus (plural: menisci). A meniscus forms at most air–water interfaces—including those found in narrow tubes, such as xylem. Menisci form because adhesion and cohesion pull water molecules up along the sides of the tube, while gravity pulls the water surface down in the middle.
➢ Surface tension is a force that exists among water molecules at an air–water interface. In the body of a water column, all the water molecules are surrounded by other water molecules and form hydrogen bonds in all directions. Water molecules at the surface, however, can form hydrogen bonds only with the water molecules beside and below them. Because they have fewer neighbors nearby, surface molecules share stronger attractive forces and bind together more tightly. This enhanced attraction results in tension that minimizes the total surface area.
■ Objections to Capillary Force Theory:
(i) Like root pressure, capillary action can transport water only a limited distance. This process moves water along the surfaces of mosses and other low-growing, non-vascular plants, but it can raise the water in the xylem of a vertical stem only about 1 meter with the normal diameter (0.03 mm).
Thus, root pressure and capillary action cannot explain how water moves from soil to the top of a redwood tree that can grow 5 to 6 stories high.
(ii) In the plants, where only tracheids are present, capillarity cannot operate due to the presence of end walls in tracheids. Vessels are not found in gymnosperms whereas these trees are enormously long.
(iii) Soil water is not directly connected with the vessels, hence this theory cannot be functional in vessels-bearing plants.
(iv) Force of capillary can operate only when one end of a narrow tube dips in water. In roots, the xylem channels are not in direct contact with soil water.
(v) Capillarity operates easily in plants having narrower vessels but tall plants have rarely such vessels.
04. COHESION-TENSION THEORY / TRANSPIRATION PULL THEORY
The leading hypothesis to explain long-distance water movement in vascular plants is the cohesion-tension theory, which states that water is pulled from roots to the tops of trees along a water potential gradient, via forces generated by transpiration at leaf surfaces. This process relies on two of the forces involved in capillary action, namely, cohesion and tension.
This theory was proposed by Dixon and Jolly in 1894. This is the most accepted theory of water movement through plants. This was further improved by Dixon in 1914. This theory is based on following steps:
● Step 1. Water vapor diffuses out of leaf: Water vapor diffuses through the stoma to the atmosphere.
●Step 2. Water evaporates inside leaf: Water is lost from mesophyll cells to the intercellular spaces as a result of transpiration. As water exits the leaf, the humidity of the spaces inside the leaf drops, causing water to evaporate from the menisci that exist at the air–water interfaces.
● Step 3. Water is pulled out of xylem: Due to loss of water from mesophyll cells, the DPD (diffusion pressure deficit) increases. The resulting tension created at the menisci pulls water that surrounds nearby cells, which in turn pulls water out of the xylem.
● Step 4. Water is pulled up xylem: Tension is transmitted from water in leaf xylem through the stem all the way to root xylem by cohesion (continuous hydrogen bonding).
● Step 5. Water is pulled out of root cortex: Water moves by osmosis as a result of the active transport of ions into the xylem.
● Step 6. Water is pulled from soil into root: Water enters the root by osmosis as a result of the active uptake of ions into root cells.
The transmission of pulling force from the leaf surface to the root is possible because (i) xylem forms a continuous network from roots to leaves, so there are continuous columns of water throughout the plant, and (ii) all of the water molecules are bonded to each other through hydrogen bonds (cohesion).
Note that the plant does not expend energy to create the pulling force. The force is generated by energy from the Sun, which drives evaporation from the leaf surface. Water transport is solar powered.
In effect, the cohesion-tension theory of water movement states that, because of the hydrogen bonding between water molecules, water is pulled up through xylem in continuous columns. (Ref: Biological Science, Sixth Edition, Scott Freeman et. al., 2017).
■ Evidences in Support of the Theory
Following evidences have been put forth to support this theory:
1. The osmotic pressure of mesophyll cells has been recorded up to 20 atm which is sufficient for ascent of sap.
2. It is purely a physical process which does not require metabolic energy.
3. The tensile strength of xylem is in between 20 – 300 atm which is enough to prevent any breaking of water column.
4. The theory gets further support when it is compared with the porous pot experiment. When at the top of a continuous water column a porous pot is placed and its water is subjected to evaporation, the water column, is put under tension. Due to cohesive property of water molecules, this column does not get broken while evaporation continues. It is said that a similar kind of mechanism operates in plants.
■ Objection to Cohesion-Tension and Transpiration Pull Theory
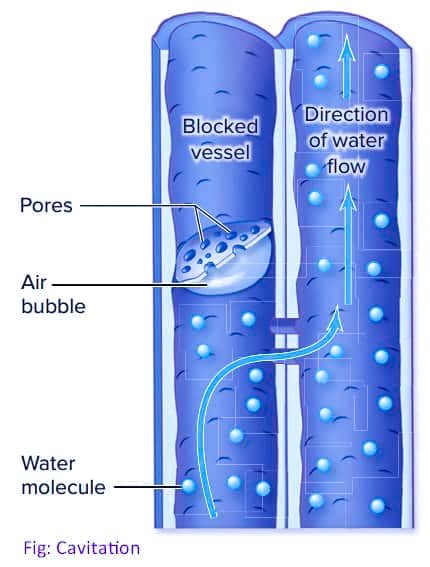
A serious objection raised against this theory was that the gas dissolved in water formed bubbles under tension and high temperature. The presence of such an air bubble would break the continuity of water column and thereby stop ascent of sap due to transpiration pull. A gas-filled bubble can expand and block the tracheid or vessel, a process called cavitation. Cavitation stops water transport and can lead to dehydration and death of part or all of a plant.
But Scholander et.al. (1957) proposed that the continuity of water column is maintained because of the presence of pits in the lateral walls of tracheids or vessels, and air bubbles are generally larger than these openings. In this way, bubbles cannot pass through the pits to further block transport. Freezing, drought conditions, or deformation of cells can also cause small bubbles of air to form within xylem cells, especially with seasonal temperature changes. Cavitation is one reason older xylem often stops conducting water.
{kind=link}
[/su_note]
MINERAL TRANSPORT
► UPTAKE OF MINERAL IONS
Unlike water, all the minerals cannot be absorbed passively because of two reasons:
01. Minerals are present in soil as ions and ions cannot move across the cell membranes.
02. Concentration of minerals inside root cells is much higher than in soil water.
Therefore, mineral ions move through the plasma membrane of root hair cells by active transport mediated by carrier protein molecules, also called active transport molecules. Their proteins are located in the plasma membrane of root hair cells. The energy needed for their active transport is supplied by ATP through the action of enzyme ATPase also present on the plasma membrane of epidermal cells of root hairs. These establish an electrostatic proton gradient for energy supply. Therefore, the movement of ions of solutes from epidermal cells through cortical cells into the cells of endodermis is along the concentration gradient.
The endodermal cells have many transport proteins in their cell membrane. Due to the presence of suberin in their wall, they allow movement of certain ions inwards, into the xylem but not outwards. Different ions are transported at varying rates. The transport proteins of endodermal cells act as controls to adjust the quantity and types of ions, that reach the xylem. The inward flow of mineral ions from endodermal cells to the xylem is also along the concentration gradient. In xylem vessels, they move along with the flow of water.
► TRANSLOCATION OF MINERAL IONS
Once the ions absorbed from the soil reach xylem through passive or active uptake or a combination of the two, they are further transported up the stem to all parts of the plant through the transpiration stream.
Analysis of xylem exudates reveals that though some of the nitrogen travels as inorganic ions, much of it is carried in the organic form as amino acids and related compounds. Similarly, small amounts of phosphorus and sulphur are also carried as organic compounds. Some exchange of materials also takes place between the xylem and phloem.
The chief sinks for the mineral elements are the growing regions of the plant such as the apical and lateral meristems, young leaves, developing flowers, fruits and seeds, and the storage organs. Mineral ions are unloaded at the fine ending of veins through diffusion and active uptake by the cells.
Remobilisation of mineral ions particularly from older senescing parts takes place to younger organs. The older dying leaves export much of their mineral contents to younger leaves. Similarly, before the leaf fall in deciduous plants, minerals are removed to other parts. Phosphorus, sulphur, nitrogen, and potassium are the elements that are the most readily mobilized. Elements that are structural components like calcium are not remobilized.
[/su_note]
PHLOEM TRANSPORT
Most carbohydrates are manufactured in leaves and other green parts as a result of photosynthesis and are distributed through the phloem to the rest of the plant. This process, known as translocation, provides suitable carbohydrate building blocks for the roots and other actively growing regions of the plant.
Translocation refers to the movement of sugars by bulk flow in multiple directions throughout a plant— but specifically, from sources to sinks. In vascular plants, a source is a tissue where sugar enters the phloem (leaves, region of photosynthesis and storage); a sink is a tissue where sugar exits the phloem (the region of utilization).
As the source-sink relationship is variable, the direction of movement of organic solutes in phloem can be bidirectional i.e., upwards or downwards; or multidirectional i.e., upward, downward and lateral translocation. This contrasts with that of the xylem where the water movement is always unidirectional, i.e., upwards.
The phloem sap that flows through vascular tissue is often dominated by the disaccharide sucrose—table sugar. Phloem sap can contain small amounts of minerals, amino acids, mRNAs, hormones, and other compounds as well.
■ Direction of Translocation of Solutes
Solutes are translocated in the plants in downward, upward or radial direction. Path of upward and downward translocation is the phloem, while the path of radial translocation is medullary rays.
➢ Downward translocation: It is of the most important type, i.e., from leaves to stem and roots.
➢ Upward translocation: From leaves to developing flowers, buds, fruits and also during germination of seeds and
tubers, etc.
➢ Radial translocation: From pith to cortex and epidermis.
■ Evidences in Favour of Downward Conduction of Solutes
Downward conduction of solutes that takes place through phloem is evident from the following points.
► Suitability: Phloem has long tubes (most suitable for long distance transport) placed one above the other. Thus, phloem seems to be the only suitable tissue for the downward conduction of solutes.
►Chemical analysis of phloem sap: Chemical analysis of sieve tube sap proves that concentrated solution of sucrose is translocated from the place of synthesis to other parts of the plant body. Glucose and fructose are sometimes found in traces only. The amount of sucrose is more in phloem sap during the day and less at the night.
► Isotopic studies: The movement of sugars and other substances can be followed in phloem using radioactive labels. Radioactive carbon dioxide (14CO2) can be incorporated into glucose as a result of photosynthesis. Glucose molecules are used to make the disaccharide sucrose, which is transported in the phloem. Such studies have shown that sucrose moves both up and down in the phloem.
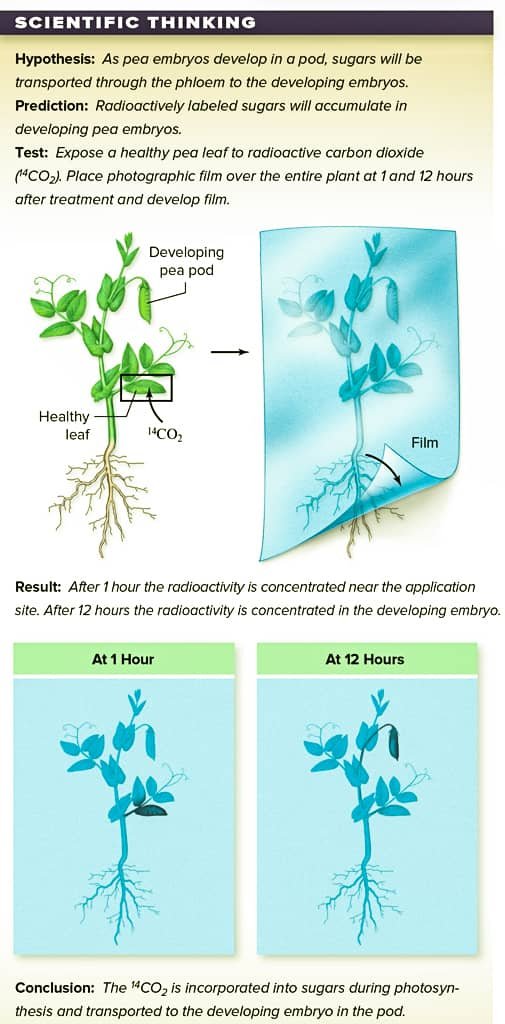{kind=link}
► Ringing or girdling experiment: In this experiment, all the tissues of the stem outside the vascular cambium are removed in a ring. The upper part of the plant is attached to the lower part only by an external xylem cylinder and pith. Accumulation of food occurs in the form of swelling just above the ring, which suggests that in absence of phloem, downward translocation of food is stopped. In a girdled plant, roots die first and ultimately shoot dies. This is because the upper part of stem gets ample amount of water supply (as transport of water and minerals occurs through the xylem). But as the roots die due to starvation, the upper part of the stem also dies in the course of time.
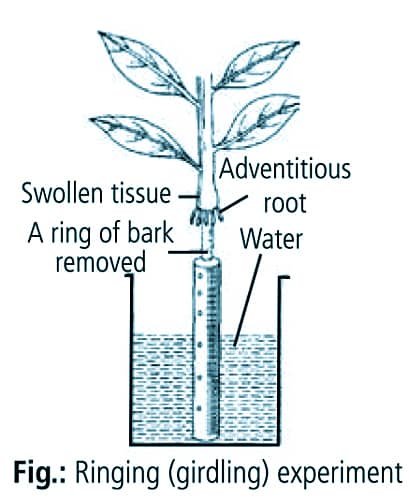{kind=link}
■ Anatomy of Pholem
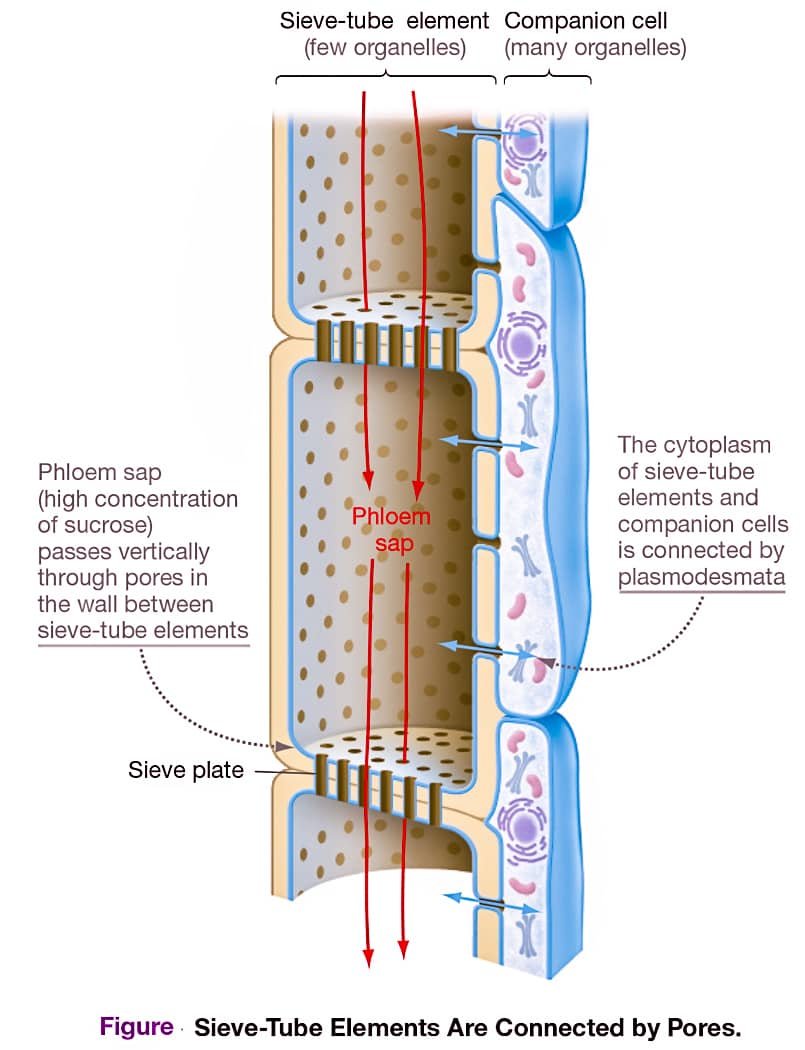{kind=link}
Phloem consists largely of two cell types, sieve-tube elements and companion cells. Unlike the tracheids and vessel elements that make up most of the xylem, sieve-tube elements and companion cells are alive at maturity and lack secondary cell walls.
In most plants, sieve-tube elements lack nuclei and most other organelles. They are connected to one another, end to end, by perforated sieve plates. The pores, which are simply enlarged plasmodesmata, create a direct connection between the cytoplasms of adjacent cells. They are responsible for transporting sugars and other nutrients. Companion cells, in contrast, have nuclei and a rich assortment of ribosomes, mitochondria, and other organelles. Companion cells are located adjacent to sieve-tube elements and maintain the cytoplasm and plasma membrane of sieve-tube elements i.e., they are “support staff.”
The sieve-tube elements in the phloem represent a continuous system for transporting sugar throughout the plant body.
Vascular bundles, which contain xylem and phloem, run the length of plants, and certain bundles extend into specific branches, leaves, and lateral roots. Phloem sap does not move from one vascular bundle to another—instead, each bundle is independent.
Based on these structural features, the physical relationships observed between sources and sinks in herbaceous plants are logical. For example, the phloem in the leaves on one side of a plant connects directly with the phloem of branches, stems, and roots on the same side, through a specific set of vascular bundles.
The phloem sap contains disaccharide sucrose (=table sugar), small amounts of minerals, amino acids, mRNAs, hormones, and other compounds as well.
■ Mechanism of Translocation of Solutes by Pholem
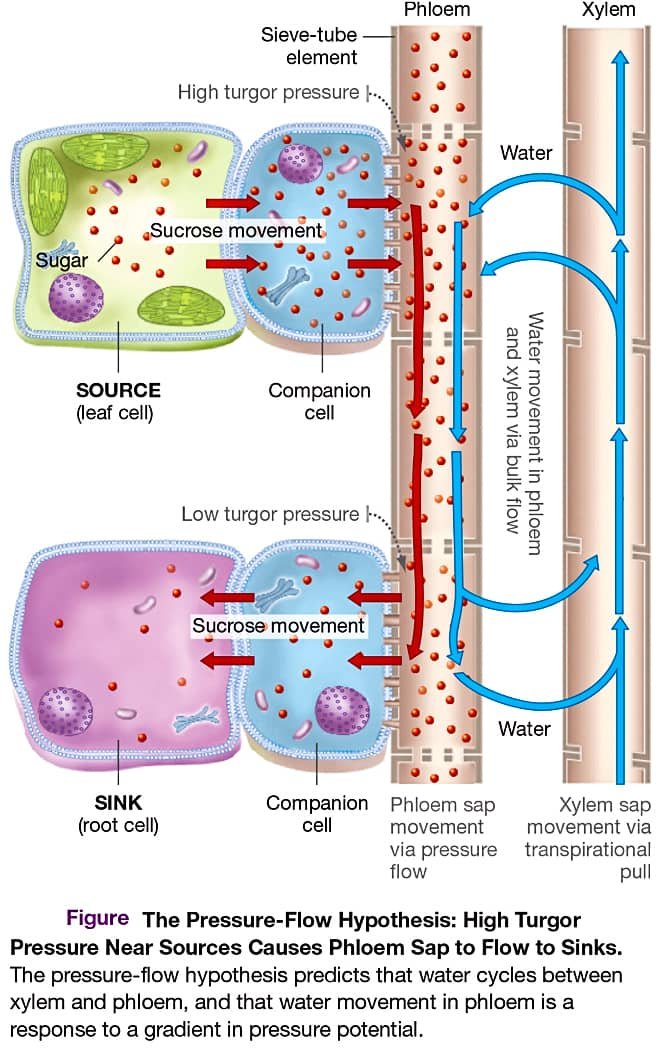{kind=link}
The most widely accepted model of carbohydrate transport through the phloem is the pressure–flow hypothesis. Dissolved carbohydrates flow from a source (such as the mesophyll of leaves) to a sink (such as the cortex of roots, growing tips of roots and stems, and in developing fruits) where they are unloaded and used. Sources and sinks can change across the life of a plant. As needs change, the direction of phloem flow can change.
According to this theory, translocation of solutes takes place in bulk, through phloem along a gradient of turgor pressure from the higher concentration of its soluble form (source) to lower concentration of its soluble form (sink).
Because in leaves (source), the osmotic concentration remains always high (due to photosynthesis) and in roots (sink) osmotic concentration remains low, so mass flow of organic food continues from leaves to roots (i.e., along a gradient of turgor pressure).
In a process known as phloem loading, carbohydrates (mostly sucrose) present in mesophyll cells are entered into the sieve tubes through their companion cells by an active process. Some sucrose travels from mesophyll cells to the companion and sieve cells via the symplast. Much of the sucrose arrives at the sieve cell through apoplastic transport and is moved across the membrane via a sucrose and H+ symporter. This energy-requiring step is driven by a proton pump. Companion cells and parenchyma cells adjacent to the sieve tubes provide the ATP energy to drive this transport. Unlike vessels and tracheids, sieve cells must be alive to participate in active transport.
A high osomotic concentration, therefore, develops in the sieve tubes of the source. Thus, the sieve tubes absorb water from the surrounding xylem by osmosis and develop a high turgor pressure. It causes the flow of organic solution towards the area of low turgor pressure. Low turgor pressure is maintained in the sink region by converting soluble organic substances into insoluble forms. Most of the water at the sink then diffuses back into the xylem, where it may be either recirculated or lost through transpiration.
Transport of sucrose and other carbohydrates within sieve tubes does not require energy. But the pressure needed to drive the movement is created through energy-dependent loading and unloading of these substances from the sieve tubes.
● Objections to this theory are as follows:
– Vacuoles of the adjacent sieve tube cells are not continuous. The cytoplasm present near the sieve plates exerts resistance to the mass flow.
– Catalado et al (1972) have observed the rate of flow of water (72 cm/hr) and solutes (35 cm/hr) to be different in the same sieve tube.
– Phloem transport is not influenced by water deficit.
– The cells at the source end of mass flow should be turgid but they are often found to be flaccid in case of germinating tubers, corms, etc.
● Factors affecting translocation of solutes:
Various factors affecting the translocation of solutes have been summarized in the given flow chart.
{kind=link}