













In this tutorial, we have discussed ‘what is transpiration in plants‘, ‘type of transpiration‘, ‘Mechanism of transpiration‘ etc.
TABLE OF CONTENTS
Transpiration Definition
Plants absorb a large quantity of water from the soil. Only a small part of it (1-2%) is utilised by the plant for building up the processes, while bulk of the water (98–99%) is lost into the air from the leaves and other aerial parts of the plant. This loss of water in the form of vapours from the aerial parts of the plant is called transpiration.
The amount of water lost by transpiration is enormous, i.e., 2–3 litres per day in sunflower plant, 35–45 litres in apple and up to 1 tonne per day in elm tree. It is estimated that a common plant loses 98–99 per cent of the water through transpiration.
► Read More: Different Types of Fruits in Biology & its Importance ► Read More: Why Seed Dispersal is important? Agents of Seed Dispersal ► Read More: What is Pollination? Definition, Types, Agents, Significance ► Read More: Reproduction in Plants : Sexual and Asexual reproduction ► Read More: Stem Anatomy || Monocot and Dicot Stem Cross Section || ► Read More: Inflorescence Types || Racemose, Cymose, Mixed, Specialized
Demonstration of Transpiration
Take a small, healthy and well-watered potted plant. Cover the pot and the soil with polythene. Place the pot over a dry glass plate near a window. Cover the pot with a dry bell jar. Seal the edges of bell jar with grease or vaseline. Leave the apparatus undisturbed for some time. Soon the water drops will be observed on the inner surface of the jar. These drops have been produced due to transpiration.

TABLE: Differences between Transpiration and Evaporation
| Sl. No. | Transpiration | Evaporation |
| 01. | It is the loss of water from the free surface of the cells. | 1. It is the loss from the free surface of water. |
| 02. | It is a physiological process. | It is purely a physical process. |
| 03. | It involves living tissues. | It involves nonliving matter. |
| 04. | It is controlled by processes like osmotic potential and water potential. | It is not controlled by any such process. |
| 05. | Light, CO2 concentration, pH and hormones regulate and control transpiration. | Light, CO2 concentration, pH and hormones do not influence evaporation. |
| 06. | It is comparatively a slow process. | It is faster. |
| 07. | It is influenced by the anatomy of the transpiring organ. | There is no such influence. |
| 08. | It is regulated by the amount of water available to the plant. | It goes on as long as the water is present. |
TABLE: Differences between Transpiration and Guttation
| Sl. No. | Transpiration | Guttation |
| 01. | It occurs mostly during the day when the temperature is comparatively higher. | It occurs during the cold hours of night and early morning. |
| 02. | Water is lost in the form of vapours. | Water is lost in the form of liquid droplets. |
| 03. | It occurs through stomata, lenticels and cuticles. | It occurs through hydathodes. |
| 04. | It occurs through all aerial plant parts. | It occurs at vein-ends on the margins or tips of the leaves where hydathodes are present. |
| 05. | It takes place under dry conditions. | It takes place under humid conditions. |
| 06. | Excessive transpiration under a deficient water supply can cause wilting. | It does not take place under a deficit water supply and never results in wilting. |
| 07. | It takes place when stomata are open. | It takes place only under conditions of adequate water supply and cold and humid hours of the night or early morning. |
[/su_table]
Types of Transpiration
Most of the transpiration occurs through foliar organs or the surface of the leaves. It is known as foliar transpiration. It accounts for over 90% of the total transpiration. Transpiration also occurs in young stems, flowers, fruits, etc. Mature stems transpire very little. Depending upon the plant surface, transpiration is of three types stomatal, cuticular and lenticular.
1. Stomatal Transpiration
Stomata are minute pores in the epidermis. Their opening and closing are controlled by guard cells. Maximum loss of water vapours takes place through these pores. The loss of water vapours through stomata amounts to about 90% of the total water loss.
Stomata are mostly situated on leaves but in green stem also, they are found in the epidermis as well. The number of stomata is normally more on the lower side than on the upper side of the leaf. In xerophytes, stomata are confined to the lower side. In submerged plants, they are absent but in floating plants, they are present on the upper side of leaves.
In dorsiventral leaves, the lower surface has a large number of stomata. Therefore, it transpires more vigorously than the upper surface. In isobilateral leaves, stomata are uniformly distributed on both surfaces. Thus in such leaves, transpiration is more or less equal from both the surfaces.

2. Cuticular Transpiration
The cuticle is a layer of wax-like covering on the epidermis of leaves and green stems. The cuticle is impervious to water, so the water lost through it is less. Up to 10% of the total transpiration may take place through it. With the increase in thickness of the cuticle, the loss of water vapour through it is reduced.
3. Lenticular Transpiration
Loss of water vapour also takes place through lenticels of woody stems and is called lenticular transpiration, Lenticular transpiration is only 0.17% of the total transpiration. It continues day and night because lenticels have no mechanism of closure.
Take small pieces of filter paper and dip them in 3% cobalt chloride solution. Take them out, squeeze the excess solution and dry them in an oven or desiccator. Dry cobalt chloride papers appear blue in colour. Take a potted plant with dorsiventral leaves. Select a healthy leaf and clean its two surfaces with dry cotton. Place cobalt chloride blue paper on each of its two surfaces. Immediately, cover them with two glass slides with vaselined margins supported by leaf clasp. Watch the colour of the two cobalt chloride papers. Within a few minutes, cobalt chloride paper on the lower surface turns pink. Thereafter, cobalt chloride paper of the upper surface also becomes pink. The dry cobalt chloride paper turns pink when it becomes moist. Cobalt chloride papers of both surfaces turn pink. This shows that both the surfaces are transpiring and losing water. In the dorsiventral leaf, stomata are generally present only on the lower surface. The lower surface is transpiring through stomata and the upper surface is performing only cuticular transpiration. It further shows that stomatal transpiration is more rapid than cuticular transpiration.
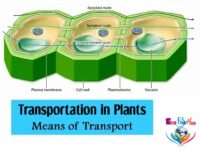
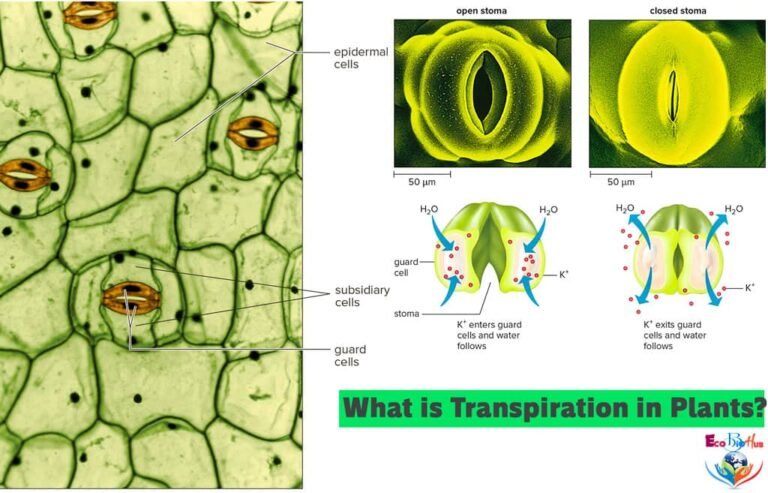
Structure of the Stomatal Apparatus
The stomata (singular: stoma) are the only apertures connecting the outer atmosphere with the intercellular spaces of the leaf. Thus, they are the main pathways through which water vapour diffuses to the outer atmosphere during transpiration. The average size of a typical stomatal pore varies from 3–12 μm in width and 10–14 μm in length. Stomatal width increases when they are close.
Each stoma is an elliptical pore found in the epidermis of leaves and green stems. The pore is bounded by two kidney-shaped epidermal cells called guard cells. The guard cells control the opening and closing of stomata, which, in turn, regulate the loss of water from the leaves to the atmosphere. The wall of guard cells towards the stoma is thickened while the outer wall is thin. The guard cells differ from ordinary epidermal cells not only in shape but also in having chloroplasts.
In dicotyledons, the inner walls of the guard cells surrounding the stoma are thicker than the outer walls. The inflow of water in guard cells causes bulging of the outer walls, drawing the inner walls apart and opening the pore. Decrease in turgor pressure of the guard cells causes the inner walls to return to their original position resulting in the closure of the pore.
In monocotyledons, guard cells are dumbbell-shaped with thick intervening walls. The flow of water increases the turgidity of the guard cells. The regions with thin walls bulge drawing the thick walls apart and opening the pore. In Cyperus, the guard cells are both kidney-shaped and dumb-bell-shaped.
Stomata are found on the lower (abaxial) as well as upper (adaxial) surfaces of the leaf. However, the number of stomata at the upper and lower surfaces of leaves varies among species. In monocot leaves, the number of stomata is almost the same on both surfaces. But, leaves of dicot plants usually have fewer stomata on the upper surface.

Based on stomatal distribution, plants are divided into categories: apple and mulberry type, potato type, oat type, water lily type, potamogeton type.
| Type of plant | Type of stomata | Location of stomata |
| Apple and mulberry type | Hypostomatous | On the lower surface of the leaf, e.g., apple, peach. |
| Potato type | Amphistomatous | On both surfaces of leaf but numerous on the lower surface, e.g., potato, bean. |
| Oat type | Isostomatous | Equally distributed on both surfaces of the leaf, e.g., wheat, rice, potato, bean. |
| Waterlily type | Epistomatous | Only on the upper surface of the leaf. These are found in plants with floating leaves. |
| Potamogeton type | Astomatous | Either absent or non-functional. This is the characteristic feature of submerged hydrophytes. |
Mechanism of Transpiration
Spongy mesophyll in the leaves has large air cavities which communicate to the outside through stomata. The mesophyll cells maintain their turgidity by absorption of water from the xylem of leaf bundles. The air inside the air cavities becomes saturated with water vapour due to the evaporation of water from the moist cell walls. The energy for it is obtained from the radiant energy of the sun during the day and metabolic energy from respiring tissues during the night.
The water potential of saturated air within the leaf is high and of unsaturated air outside the leaf is low. Thus, water vapours from the air cavities move to the outside air through the stomatal opening. This lowers the water potential of air chambers in the mesophyll and more water is drawn from the mesophyll cells. This continues till the stomata remain open.
Stomatal index may be defined as the percentage number of stomata as compared to all the epidermal cells (including stomata) in a unit area of the leaf. Salisbury (1927) gave a correlation between the number of stomata and the epidermal cells per unit area: where I = stomatal index, S = number of stomata per unit area, E = epidermal cells Stomatal index is independent of environmental conditions and not affected by conditions like age of the plant and position of the leaf. Note: Water potential is measured in bars, atmospheres and megapascals. No water movement occurs in a cell, when OP = TP Manometer: An instrument used for measuring pressure or tension in liquids and gases. Depending upon the periods of opening and closing stomata are of four types as given below: ➢ Scotoactive stomata are stomata that open in dark and close during the daytime. These occur in succulents. ➢ Both red and blue lights stimulate stomatal opening though blue light is slightly more effective. ➢ In dry weather, stomata tend to close while in a humid environment they remain open for a longer period. ➢ Rise in temperature induces stomatal opening and vice versa. ➢ At 38°- 40°, stomata open even in dark. Following are the theories of stomatal movement: This hypothesis was given by Schwendener (1881). According to this theory, chloroplasts of guard cells perform photosynthesis in light and produce sugar. Sugar increases the osmotic concentration of guard cells. Guard cells absorb water, swell up and bend outwards creating a pore in between. However, the photosynthetic activity of chloroplasts of guard cells seems to be negligible. Therefore, this hypothesis is not acceptable. Objections – Photosynthetic activity of the guard cell chloroplasts is not confirmed. – Sugar does not occur in detectable quantity in the guard cells. – In some plants stomata open during night. According to Llyod (1908), the conversion of starch into sugar in the day time and vice versa at night causes the opening and closing of stomata. Later, Sayre (1923) proposed the Starch-Sugar hypothesis and this was modified by Steward (1964). According to this hypothesis, the turgidity of guard cells is responsible for the opening and closing of stomata. The turgidity of guard cells is controlled by a change in the osmotic potential caused by the interconversion of starch stored in guard cells into glucose. Hydrolysis of starch into glucose is induced by the increase in pH of guard cells. (a) In Light: According to the starch-sugar hypothesis, the guard cells in the morning have a high concentration of CO2 which gets accumulated from respiration during the night. Due to the accumulation of CO2, the free H+ ions are lowered in the guard cells. This increases pH, which, in turn, induces enzyme phosphorylase. This enzyme hydrolyses starch into glucose-1-phosphate. This is changed to glucose-6-phosphate which is hydrolysed into glucose and inorganic phosphate. According to Steward, at higher pH, the opening of the stomata is caused in the following manner: (b) In Dark: In the evening, when photosynthesis stops, the closure of stomata is brought about by increased carbon dioxide contents of the leaf. It decreases pH of guard cells and glucose is phosphorylated to glucose-1-phosphate in the presence of enzyme hexokinase. In the presence of phosphorylase, glucose-1-phosphate is changed into starch. This results in the fall of the osmotic concentration of guard cells. They lose water to adjacent cells and turgidity. Due to the loss of turgidity, guard cells shrink and close the pore in between them. Objections (a) Glucose has not been detected in the guard cells of open stomata. (b) In the presence of light when starch disappears from guard cells, malic acid appears but not sugars. (c) Starch has not been reported in the guard cells of many monocots (e.g. onion). (d) According to this theory, OP of guard cells is increased due to the formation of glucose1-phosphate in guard cells, but it is found that the presence of phosphate causes the development of the same OP as does the presence of glucose-1-phosphate. 
Sl. No.
Type of stomata
Nature of stomata
01.
Barley or cereal type (dumbbell shaped), e.g., maize, wheat.
Open for a few hours during the day.
02.
In Alfafa or leucerne type, e.g., pea, bean, grape.
Remain open throughout the day, close at night.
03.
In Potato type, e.g., potato, Cucurbita, bananas.
Open throughout day and night but close for a few hours in case of water deficiency.
04.
In Equisetum type, e.g., emergent hydrophytes.
Remain open and seldom close.
Mechanism of Stomatal Movement
1. Hypothesis of Guard Cells Photosynthesis

2. Starch-Sugar Hypothesis (Classical hypothesis)




(e) Enzyme phosphorylase helps in the conversion of starch to glucose1-phosphate but not in the formation of starch from glucose-1-phosphate.
(f) Although the relative effectiveness of different wavelengths of light is, in general, the same for photosynthesis, but for stomatal opening, blue light appears to be more effective because of its effect on permeability.
(g) While stomata open in very dim sunlight (50–100 lux) or even in the moonlight, not much photosynthesis can take place at this low intensity.
3. Malate or K+ ion Pump Hypothesis
This theory was postulated by Levitt (1974). According to this theory, the changes in turgor pressure that open and close the stomata result from the reversible intake and loss of potassium ions. It has now been established that the opening of stomata induced by light is accompanied by the transport of K+ ions from surrounding epidermal cells into the guard cells. The stomatal opening is an energy requiring active process whereas the stomatal closure is not.
The potassium ions play a three-fold role in the opening of stomata:
(i). K+ ions help in transport of maximum H+ into the chloroplast.
(ii). K+ ions along with organic acid raise the osmotic potential of guard cells to cause stomatal opening.
Active K+ transport mechanism can be explained in the following steps:

(a) In Light: When a leaf is exposed to light, the starch of the guard cells is converted into malic acid. Malic acid dissociates into malate and H+ ions. H+ ions are transported to epidermal cells and K+ ions enter the guard cells. This process is called ion exchange. K+ ions are balanced by organic ions (malate). Some Cl– ions are also taken to balance the K+ ions. H+– K+ ions exchange is an active process and needs energy (ATP). This is supplied either by respiration or by photosynthesis. Increased percentage of K+ and malate ions in the guard cells increases their osmotic pressure and more water enters from the surrounding cells by endosmosis.
(b) In Dark: Photosynthesis stops but respiration continues. This increases CO2 concentration in the substomatal cavities. Due to a higher concentration of CO2, in substomatal cavities, active transport of K+ ions into guard cells ceases (Sharpe and Zeiger, 1981). The formation of abscisic acid promotes the reversal of H+– K+ pump (Cowen, 1982). Malate ions present in the guard cells cytoplasm produce malic acid with H+ ions. Malic acid inhibits its own synthesis by lowering the activity of PEP carboxylase. These changes induce reversal of ion movements and K+ ions are transported out from guard cells into surrounding cells. This decreases OP of guard cells, thereby causing the movement of water into the surrounding cells. Gaurd cells become flaccid and the stomatal pore is closed.

Noggle and Fritz (1976) have summarized the sequence of events occurring during light-induced stomatal opening as follows:

► Factors Affecting the Opening and Closing of Stomata
Following factors affect the opening and closing of stomata:
1. Light: In most of the plants, stomata open during the day and close at night. Light acts in two ways:
(a) It produces sugars through photosynthesis.
(b) It decreases CO2 concentration, thus, inducing malic acid synthesis. Malic acid is involved in H+ ⇌ K+ exchange mechanism.
Stomatal opening early in the day is associated with a peak of K+ and is thought to be triggered by blue light. Later in the day, photosynthetically active light stimulates and maintains stomatal opening through the synthesis and accumulation of sucrose.

2. Abscisic acid (ABA) Hormone: Formation of ABA induces closing of stomata. This has an adaptive value to prevent excessive water loss. The scarcity of water or intense solar radiation induces the synthesis in plants.

3. Soil moisture: Stomata close when the soil is dry and root cells experience water stress. ABA is synthesized
by root cells and is quickly translocated to the leaves through xylem where it induces stomatal closure.
4. Intracellular CO2 Concentration:
A high level of CO2 concentration causes stomatal closure. Mechanism of stomatal closure is similar to that of ABA. It triggers the opening of Cl- channels, leading to the efflux of Cl- ions and is followed by membrane depolarization. Depolarization, in turn, triggers the opening of outwardly rectifying K+ channels and subsequent K+ efflux.
Low CO2 concentration increases the pH which induces the synthesis of malic acid. This causes the opening of stomata.
5. Atmospheric Humidity: In a humid environment, the stomata remain open for longer periods while in a dry environment, they remain closed for longer periods.
6. Temperature: A rise in temperature stimulates the activity of hydrolytic enzymes, thus accelerating the opening of stomata.
7. H+ Concentration: The rise in pH stimulates the opening of stomata and low pH favours the closing of stomata.
8. Minerals: Stomatal opening depends upon the availability of K+ ions from adjacent epidermal cells. Minerals like P, N, Mg, Ca, etc., are also essential for stomatal opening.
9. Oxygen and respiratory inhibitors: S Anaerobiosis will prevent stomatal opening. Many respiratory inhibitors such as phenolic compounds, azide, and arsenate also inhibit stomatal opening.
Significance of Transpiration
A. Advantages of Transpiration
Water is of vital importance to the plant and yet out of the total amount of water absorbed, 99 percent is lost in the form of water vapour. Transpiration has been described as a necessary evil.
Transpiration is regarded as being advantageous to the plant in many ways:
1. Transport of Minerals and Water: Transpiration helps in the rapid translocation of minerals and water through the xylem, once they enter the plant through root hairs.
2. Ascent of Sap: According to some, transpiration develops a suction force that helps in the ascent of sap.
3. Regulation of temperature: Transpiration helps in keeping the temperature of the plant low even when the plant is exposed to bright sunlight. Through its effect on water relations of plant cells and tissues, transpiration has significant indirect effects on the metabolic processes of the plant.
4. Concentration of Cell Sap: Transpiration secures the concentration of the cell sap and this helps in osmosis.
5. Development of Mechanical Tissue: Plants showing high transpiration rates exhibit adequate development of mechanical tissues, making them more tough and resistant to mechanical injury by wind, rain and pathogens like fungi and bacteria. The cell walls become thick and highly cutinized thereby increasing the resistance of plants
to pathogenic fungi and bacteria.
6. Quality of Fruits: High rate of transpiration leads to the development of fruits with increased sugar and mineral contents.
7. Roots: For proper anchoring of the plant, roots must be well-developed. The higher rate of transpiration may lead to well-developed root system.
B. Disadvantages of Transpiration
1. Expenditure of Energy: Since most of the water absorbed by the plants is transpired, transpiration involves a wasteful expenditure of energy
2. Wilting: Excessive transpiration may cause water deficit leading to wilting. Temporary wilting greatly reduces the yield of crop plants by retarding the metabolic processes. Wilting, if not recovered, may lead to the death of plants.
3. Stunted Growth: Excessive transpiration leads to stunted growth of plants.
4. Shedding of Leaves: Underwater deficit conditions, excessive transpiration leads to the production of abscisic acid which results in premature fall of leaves.
5. Reduction in Yield: Excessive transpiration and reduced availability of water check the growth of seeds and fruits.
In spite of various disadvantages, the plant cannot avoid transpiration due to their peculiar structure of leaves which is basically meant for gaseous exchange during respiration and photosynthesis. Therefore transpiration is also regarded as “necessary evil” by Curtis (1926) or “unavoidable evil” by Steward (1959).
Factors Affecting Rate of Transpiration
A. External Factors
1. Light: Light affects transpiration in two ways. Firstly, it increases transpiration by raising the temperature of leaves. Secondly, there is a close relationship between the opening of stomata and the presence of light. During the night, when there is no light, stomata remain closed resulting in minimum transpiration.
2. Atmospheric Humidity: Low and high humidity of air directly affect transpiration. When humidity is low, air becomes drier, receives moisture readily and increases transpiration. On the other hand, when humidity increases, the air becomes moist or saturated and receives less water vapours, thereby decreasing transpiration.
3. Air Temperature: Temperature also affects the rate of transpiration but indirectly. Transpiration also increases or decreases along with rising or fall in temperature.
4. Wind: Wind greatly affects transpiration. During high wind, the rate of transpiration is greatly enhanced as it removes water vapours transpired, thereby, not allowing the transpiring surface to become saturated.
5. Available Soil Water: Transpiration is directly linked to the availability of soil water. A high concentration of salts in the soil water reduces transpiration, probably by increasing the osmotic pressure of plant cells.
B. Internal Factors
1. Root-shoot Ratio: The rate of transpiration directly depends upon the efficiency of the absorbing system (roots) and the evaporating system (leaves). The rate of transpiration increases with an increase in the root-shoot ratio.
2. Leaf Area: There seems to be no relationship between the leaf area and water loss. In large plants, the transpiration rate may be more per plant though on a unit area basis, transpiration is more in the smaller plants than the larger ones.
3. Leaf Structure: The plants growing in dry conditions or arid areas show the following anatomical modifications to reduce the rate of transpiration:
(a) Presence of thick cuticle on the surface of aerial parts of the plant.
(b) Presence of a waxy coating or suberin and epidermal hairs on the surface of leaves.
(c) Reduction in the number of stomata.
(d) Presence of sunken stomata.
(e) Compact mesophyll without intercellular spaces. ,
(f) To reduce transpiration, leaves become modified into spines, thorns, scaly leaves phyllodes, etc., and the stem gets modified into phylloclade.
The rate of transpiration in a plant is an indirect measure of the rate of photosynthesis as it indicates the degree and period of stomatal opening. The amount of water consumed by a crop plant to produce a unit weight of dry matter is called water requirement of the crop. It differs from crop-to-crop. ➢ Potato, rice and sugarcane require large amounts of water. ➢ Ragi, bajra and sorghum require relatively less quantity of water. ➢ Neem, Acacia, Shisham (Dalbergla sisoo), Prosopis, etc., can grow under a low moisture supply. Coffee, tea, coconut, rosewood, teak, willows and bamboos require huge quantities of water. In our country where the soil is not well-irrigated in large tracts of land, the choice of the crop, depends largely on its water requirement. It is also of interest to note that the plants adapted to grow in arid regions also show a slow rate of vegetative growth. Anti-transpirants are chemical substances that are used to reduce the rate of transpiration without causing any adverse effect on their growth and other metabolic activities. These substances are sprayed on crop plants during the dry season to avoid wilting when the rate of transpiration is high. Anti-transpirants are of four types: These are phenyl mercuric acetate (PMA) and abscisic acid (ABA) , and those with high CO2 concentration; minimize the rate of transpiration by closure of stomata. With stomatal closure, diffusion of carbon dioxide gas into the leaf reduces, thus resulting in lower crop yields. Although use of PMA as an antitranspirant is quite common, but it is poisonous for fruits and vegetables. It also damages foliage by blocking the phosphorylation part of photosynthesis. CO2 is another effective antitranspirant. A slight increase in CO2 concentration from 0.03 to 0.05 percent brings about stomatal closure partially, but beyond that, it adversely affects the physiological processes such as photosynthesis and respiration by complete stomatal closure. Inhibitors of photosynthetic electron transport chain such as triazine, atrazine, simazine (herbicides) can also be used as antitranspirants at the lower concentrations.Generally, these types of antitranspirants are used for ornamentals, ground covers, and other such utility plants. These compounds (such as low viscosity waxes, colourless plastics, and silicone oils) form a film on the surface of leaves and thus reduce transpiration. This type of chemicals such as kaolin and lime water reflect the radiation falling on the leaves, thus decreasing their temperature. These check the water loss without any effect on carbon dioxide assimilation. Kaolin does not affect any metabolic activity and usually forms a thin white film on the leaf when sprayed (2–5%). These chemicals such as cycocel (CCC) suppress leaf area development as well as shoot growth, promote root growth, and induce stomatal closure thus making the plants drought-resistant. ► Uses of Anti-transpirants Antitranspirants are generally employed for reducing transpiration loss to horticultural plants. Several strategies to improve productivity have been adopted by farmers and plant physiologists. The work on stomatal physiology to reduce water loss by spraying antitranspirants has given encouraging results. 1. Field Crops: Usage of anti-transpirants helps in growing high yield crops with high water requirements in areas which go dry soon for most of the year. 2. Fruit Cracking: Banana, Tomato, Cherry, etc., crack due to rainfall during the later stages of their ripening. Anti-transpirants could greatly reduce water absorption of fruits and save them from cracking 3. Ornamental Horticulture: Usage of film forming anti-transpirants reduces transpiration in them and hence can withstand desiccation and delayed irrigation. 4. Usage in Lawns: Spraying of lawns by anti-transpirants could check excessive water loss when the lawn is not watered for several days. 5. The usage of anti-transpirants in cut flowers could enhance their table keeping quality and prolongs their freshness. Transpiration has more than one purpose in plants: 1. It creates a pull for absorption and transportation of water in plants. 2. It keeps the cells turgid, thus, maintaining the shape and structure of the plants. 3. By evaporative cooling, transpiration cools leaf surfaces up to 10–15°C. 4. It supplies water for photosynthesis. An actively transpiring plant continuously needs water. Photosynthesis is affected in case the availability of water to the plant is depleted by transpiration. In rainforests, humidity remains high due to the water cycle from soil to root to leaf to atmosphere and back to the soil. This water cycle maintains a continuous supply of water to leaves to carry photosynthesis. The ratio of the amount of water transpired to the amount of dry matter formed is called the transpiration ratio Plants with crassulacean metabolism (CAM) keep their stomata closed during the day: Therefore, their water demand is low. In such plants, CO2 is absorbed and stored during the night when transpiration is low. In desert plants that are adapted for CAM photosynthesis, the rate of transpiration is very low and the rate of carbon assimilation is high. So, the transpiration ratio of CAM plants is about 50. Plants obtain their carbon and most of the oxygen from CO2 present in the atmosphere. They obtain their remaining nutritional requirements from minerals and water (for hydrogen) obtained from the soil. Unlike water, all the minerals cannot be absorbed passively because of two reasons: 1. Minerals are present in the soil as ions and ions cannot move across the cell membranes. 2. Concentration of minerals inside root cells is much higher than in soil water. Therefore, mineral ions move through the plasma membrane of root hair cells by active transport mediated by carrier protein molecules, also called active transport molecules. Their proteins are located in the plasma membrane of root hair cells. The energy needed for their active transport is supplied by ATP through the action of enzyme ATPase also present on the plasma membrane of epidermal cells of root hairs. These establish an electrostatic proton gradient for energy supply. Therefore, the movement of ions of solutes from epidermal cells through cortical cells into the cells of endodermis is along the concentration gradient. The endodermal cells have many transport proteins in their cell membrane. Due to the presence of suberin in their wall, they allow movement of certain ions inwards into the xylem but not outwards. Different ions are transported at varying rates. The transport proteins of endodermal cells act as controls to adjust the quantity and types of ions, that reach the xylem. The inward flow of mineral ions from endodermal cells to the xylem is also along the concentration gradient. In xylem vessels, they move along with the flow of water. growing regions of the plant such as the The amount of water transpired by the plant divided by the amount of CO2 assimilated through photosynthesis is called Transpiration ratio. It is a parameter to assess the effectiveness of plants in balancing the loss of water coupled with adequate intake of CO2 for photosynthesis. ► Causes of high transpiration ratio (i) The concentration gradient responsible for water loss is approximately 50 times more as compared to that of the intake of CO2. This might be because of the low concentration of CO2 in the air (0.036%) and relatively high concentration of water vapour within the leaf. (ii) The diffusion rate of CO2 is about 1.6 times slower through the air than that of water. This is because the CO2 molecule being bigger than the water molecule, has a lower diffusion coefficient. (iii) In order to be absorbed in the chloroplast, CO2 must diffuse through the plasma membrane and the chloroplast membrane through the cytoplasm, which contribute to the resistance of the CO2 diffusion passage. ► Transpiration ratios of C3, C4, and CAM plants are as follows: ➢ C3 plants have a transpiration ratio of 400 (hence, water use efficiency is 0.0025 or 1/400; approximately 400 water molecules are lost for one carbon dioxide molecule assimilated through photosynthesis). ➢ C4 plants have a transpiration ratio of 150 (comparatively less water gets transpired per molecule of CO2assimilated. This is because C4 photosynthesis results in a lower CO2 concentration in the intercellular air space, thus creating a larger driving force for the uptake of CO2 and allowing these plants to operate with smaller stomatal apertures and thus lower transpiration rates. ➢ CAM plants have an even lower transpiration ratio, approximately 50. Because their stomata open at night, so transpiration is much lower at night. — Usually in plants growing in moist, warm soil and under humid conditions the droplets of water ooze out along the margin or tip of the leaf. The exudation of water as droplets from the tips and margins of leaves, forced out through leaf pores by root pressure, is called guttation. — The term guttation was coined by Bergerstein. — The factors that favour guttation are high water absorption, high root pressure and reduced or no transpiration. — Guttation is supposed to be due to positive root pressure. Normally it occurs in night. — The maximum rate of guttation is found in Colocasia nymphefolia. Guttation occurs through specialised structures (pores/opening of veins) called hydathodes or water stomata or water gland. Hydathodes are mainly found in aquatic and herbaceous plants and in grasses. Hydathode represents modified bundle-ends. Guttation liquid is not pure water but a solution containing a great number of dissolved substances.
Anti-Transpirants
1. Stomatal closing type
2. Film Forming Substances
3. Reflectance type
4. Growth retardants
Transpiration and Photosynthesis – A Compromise
Uptake of Mineral lons
Guttation
